| Вирусы иммунодефицита человека | |
|---|---|
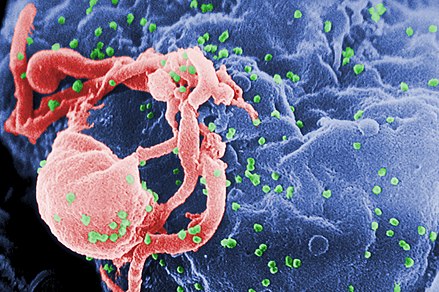 | |
| Сканирующая электронная микрофотография ВИЧ-1 (зеленого цвета), почкования из культивируемых лимфоцитов . Множественные круглые бугорки на поверхности клетки представляют собой места сборки и почкования вирионов. | |
| Научная классификация | |
| (без рейтинга): | Вирус |
| Царство : | Рибовирия |
| Королевство: | Парарнавиры |
| Тип: | Artverviricota |
| Учебный класс: | Revtraviricetes |
| Заказ: | Ортервиралес |
| Семья: | Retroviridae |
| Подсемейство: | Orthoretrovirinae |
| Род: | Лентивирус |
| Включенные группы | |
| Другие лентивирусы | |
{kind=link}
В вирусах иммунодефицита человека ( ВИЧ ) являются два видом Лентивируса (подгруппа группы ретровирусов ), поражающим человек. Со временем они вызывают синдром приобретенного иммунодефицита (СПИД) [1] [2], состояние, при котором прогрессирующий отказ иммунной системы позволяет процветать опасным для жизни оппортунистическим инфекциям и раку . [3] Без лечения средняя продолжительность жизни после заражения ВИЧ составляет от 9 до 11 лет, в зависимости от подтипа ВИЧ. [4] В большинстве случаев ВИЧ - это инфекция, передающаяся половым путем, ипроисходит при контакте с кровью , предэякулятом , семенной жидкостью и влагалищными жидкостями или при их передаче . Исследования показали (как для однополых, так и для разнополых пар), что ВИЧ не передается при половом акте без презерватива, если ВИЧ-положительный партнер имеет неизменно неопределяемую вирусную нагрузку. [5] [6] Передача инфекции не половым путем может происходить от инфицированной матери ее младенцу во время беременности , во время родов при контакте с ее кровью или вагинальной жидкостью, а также через грудное молоко . [7] [8] [9] [10] В этих жидкостях организма ВИЧ присутствует как в свободном виде.вирусные частицы и вирус в инфицированных иммунных клетках .
ВИЧ заражает жизненно важные клетки иммунной системы человека, такие как Т-хелперы (в частности, CD4 + Т-клетки), макрофаги и дендритные клетки . [11] ВИЧ-инфекция приводит к снижению уровня CD4 + Т-клеток за счет ряда механизмов, включая пироптоз абортивно инфицированных Т-клеток, [12] апоптоз неинфицированных клеток-свидетелей, [13] прямое вирусное уничтожение инфицированных клеток и уничтожение инфицированные CD4 + Т-клетки цитотоксическими лимфоцитами CD8 +, которые распознают инфицированные клетки. [14] Когда CD4+ Количество Т-клеток снижается ниже критического уровня, клеточный иммунитет теряется, и организм становится все более восприимчивым к оппортунистическим инфекциям, что приводит к развитию СПИДа.
Вирусология
Классификация
| Разновидность | Вирулентность | Инфекционность | Распространенность | Предполагаемое происхождение |
|---|---|---|---|---|
| ВИЧ-1 | Высоко | Высоко | Глобальный | Обыкновенный шимпанзе |
| ВИЧ-2 | Ниже | Низкий | Западная Африка | Закопченный мангабей |
ВИЧ является членом рода лентивирусов , [15] часть семьи Retroviridae . [16] Лентивирусы имеют много общих морфологий и биологических свойств. Многие виды заражены лентивирусами, которые обычно вызывают длительные заболевания с длительным инкубационным периодом . [17] Лентивирусов передаются в виде одноцепочечных , положительно смысле , оболочечных РНК - вирусов . При попадании в клетку-мишень геном вирусной РНК преобразуется (обратная транскрипция) в двухцепочечную ДНК кодируемым вирусом ферментом, обратной транскриптазой , которая транспортируется вместе с вирусным геномом в вирусной частице. В результате вирусная ДНК затем импортируется в ядро клетки и интегрированная в клеточную ДНК с помощью фермента кодируемого вируса, интегразов и принимающих кофакторов . [18] После интеграции вирус может стать латентным , что позволяет вирусу и его клетке-хозяину избегать обнаружения иммунной системой в течение неопределенного времени. [19]Вирус ВИЧ может оставаться в спящем состоянии в организме человека до десяти лет после первичного заражения; в этот период вирус не вызывает симптомов. Альтернативно, интегрированная вирусная ДНК может быть транскрибирована , производя новые геномы РНК и вирусные белки, используя ресурсы клетки-хозяина, которые упаковываются и высвобождаются из клетки в виде новых вирусных частиц, которые начнут цикл репликации заново.
Были охарактеризованы два типа ВИЧ: ВИЧ-1 и ВИЧ-2. ВИЧ-1 - это вирус, который был первоначально открыт и назван как вирусом, связанным с лимфаденопатией (LAV), так и человеческим Т-лимфотропным вирусом 3 (HTLV-III). ВИЧ-1 более вирулентен и более заразен, чем ВИЧ-2 [20], и является причиной большинства ВИЧ-инфекций во всем мире. Более низкая инфекционность ВИЧ-2 по сравнению с ВИЧ-1 означает, что меньше людей, подвергшихся воздействию ВИЧ-2, будут инфицированы за один контакт. Из-за относительно низкой способности к передаче ВИЧ-2 в основном ограничен Западной Африкой . [21]
Структура и геном
ВИЧ отличается по структуре от других ретровирусов. Он имеет примерно сферическую форму [22] с диаметром около 120 нм , что примерно в 60 раз меньше эритроцита . [23] Он состоит из двух копий одноцепочечной РНК с положительным смыслом, которая кодирует девять генов вируса, заключенных в конический капсид, состоящий из 2000 копий вирусного белка p24 . [24] Одноцепочечная РНК прочно связана с белками нуклеокапсида, р7 и ферментами, необходимыми для развития вириона, такими как обратная транскриптаза , протеазы , рибонуклеаза и интеграза . Матрица, состоящая из вирусного белка p17, окружает капсид, обеспечивая целостность частицы вириона. [24]
Он, в свою очередь, окружен вирусной оболочкой , которая состоит из липидного бислоя, взятого из мембраны клетки-хозяина человека, когда вновь образованная вирусная частица выходит из клетки. Вирусная оболочка содержит белки из клетки-хозяина и относительно небольшое количество копий белка оболочки ВИЧ [24], который состоит из крышки, состоящей из трех молекул, известных как гликопротеин (gp) 120 , и стержня, состоящего из трех молекул gp41 , которые закрепляют структура в вирусную оболочку. [25] [26] Белок оболочки, кодируемый геном env ВИЧ , позволяет вирусу прикрепляться к клеткам-мишеням и сливать вирусную оболочку с мишенью.клеточная мембрана, высвобождающая вирусное содержимое в клетку и запускающая инфекционный цикл. [25]
Как единственный вирусный белок на поверхности вируса, белок оболочки является основной мишенью для разработки вакцины против ВИЧ . [27] Более половины массы шипа тримерной оболочки составляют N-связанные гликаны . Плотность высока, поскольку гликаны защищают лежащий в основе вирусный белок от нейтрализации антителами. Это одна из известных молекул с наиболее плотным гликозилированием, и ее плотность достаточно высока, чтобы предотвратить нормальный процесс созревания гликанов во время биогенеза в эндоплазматическом аппарате и аппарате Гольджи. [28] [29] Таким образом, большинство гликанов застопорились как незрелые «высокоманнозные» гликаны, которые обычно не присутствуют на гликопротеинах человека, которые секретируются или присутствуют на поверхности клетки. [30]Необычный процессинг и высокая плотность означают, что почти все широко нейтрализующие антитела, которые были идентифицированы до сих пор (от подгруппы пациентов, которые были инфицированы в течение многих месяцев или лет), связываются с этими гликанами оболочки или адаптированы для борьбы с ними. [31]
Молекулярная структура вирусного спайка теперь определена с помощью рентгеновской кристаллографии [32] и криогенной электронной микроскопии . [33] Эти успехи в структурной биологии стали возможными благодаря развитию стабильных рекомбинантных форм вирусного шипа путем введения межсубъединичной дисульфидной связи и мутации изолейцина в пролин ( радикальная замена аминокислоты) в gp41. [34] Так называемые тримеры SOSIP.не только воспроизводят антигенные свойства нативного вирусного шипа, но также демонстрируют ту же степень незрелости гликанов, что и нативный вирус. [35] Рекомбинантные тримерные вирусные шипы являются многообещающими кандидатами в вакцины, поскольку они демонстрируют меньше ненейтрализующих эпитопов, чем рекомбинантный мономерный gp120, который подавляет иммунный ответ на целевые эпитопы. [36]
Геном РНК состоит как минимум из семи структурных ориентиров ( LTR , TAR , RRE , PE, SLIP, CRS и INS) и девяти генов ( gag , pol и env , tat , rev , nef , vif , vpr , vpu , и иногда десятый tev , который представляет собой слияние tat , env и rev ), кодирующего 19 белков. Три из этих генов: gag , pol и env., содержат информацию, необходимую для создания структурных белков для новых вирусных частиц. [24] Например, env кодирует белок gp160, который клеточной протеазой разрезается пополам с образованием gp120 и gp41. Шесть оставшихся генов, tat , rev , nef , vif , vpr и vpu (или vpx в случае ВИЧ-2), представляют собой регуляторные гены для белков, которые контролируют способность ВИЧ инфицировать клетки, производить новые копии вируса ( воспроизвести), или вызвать болезнь. [24]
Два белка tat (p16 и p14) являются транскрипционными трансактиваторами для промотора LTR, действующего путем связывания элемента РНК TAR. TAR также может быть преобразован в микроРНК, которые регулируют гены апоптоза ERCC1 и IER3 . [37] [38] числа оборотов белка (P19) участвует в челночные РНК из ядра и цитоплазмы путем связывания с RRE - элемент РНК. VIF белок (p23) предотвращает действие APOBEC3G (клеточный белок , который дезаминирует цитидин в уридинв одноцепочечной вирусной ДНК и / или препятствует обратной транскрипции [39] ). ВПР белок (P14) аресты деление клеток в G2 / M . NEF белок (p27) вниз регулирует CD4 (главный вирусный рецептор), а также МНС класса I и класса II , молекулы. [40] [41] [42]
Nef также взаимодействует с доменами SH3 . ВПУ белок (p16) влияет на выпуск новых вирусных частиц из инфицированных клеток. [24] Концы каждой цепи РНК ВИЧ содержат последовательность РНК, называемую длинным концевым повтором (LTR). Области в LTR действуют как переключатели для контроля производства новых вирусов и могут запускаться белками либо из ВИЧ, либо из клетки-хозяина. Элемент Psi участвует в упаковке вирусного генома и распознается белками gag и rev . Элемент SLIP (TTTTTT) Участвует в рамки считывания в кляп - пол- рамки считывания , необходимой для принятия функциональной Pol . [24]
Тропизм
Термин " вирусный тропизм" относится к типам клеток, которые заражает вирус. ВИЧ может инфицировать различные иммунные клетки, такие как CD4 + Т-клетки , макрофаги и микроглиальные клетки . Поступление ВИЧ-1 в макрофаги и CD4 + Т-клетки опосредуется посредством взаимодействия гликопротеинов оболочки вириона (gp120) с молекулой CD4 на мембране клеток-мишеней, а также с корецепторами хемокинов . [25] [43]
Макрофаги-тропные (М-тропные) штаммы ВИЧ-1, или не- синцитий -inducing штаммов (НСИ, теперь называемые вирусы R5 [44] ) используют & beta ; -chemokine рецептор, CCR5 , для ввода и, таким образом , способны к репликации в как макрофаги, так и CD4 + Т-клетки. [45] Этот корецептор CCR5 используется почти всеми первичными изолятами ВИЧ-1 независимо от вирусного генетического подтипа. Действительно, макрофаги играют ключевую роль в нескольких важных аспектах ВИЧ-инфекции. Они, по-видимому, являются первыми клетками, инфицированными ВИЧ, и, возможно, источником продукции ВИЧ, когда у пациента истощаются клетки CD4 + . Макрофаги и микроглиальные клетки - это клетки центральной нервной системы, инфицированные ВИЧ.. В миндалинах и аденоидах ВИЧ-инфицированных макрофаги сливаются в многоядерные гигантские клетки, которые производят огромное количество вируса.
Т-тропные штаммы ВИЧ-1 или штаммы, индуцирующие синцития (SI; теперь называемые вирусами X4 [44] ), реплицируются в первичных CD4 + Т-клетках, а также в макрофагах и используют для входа рецептор α- хемокина, CXCR4 . [45] [46] [47]
Считается, что двухтропные штаммы ВИЧ-1 являются переходными штаммами ВИЧ-1 и, таким образом, могут использовать как CCR5, так и CXCR4 в качестве корецепторов для проникновения вируса.
Α -chemokine SDF-1 , A - лиганд для CXCR4, подавляет репликацию Т-тропных ВИЧ-1 изолятов. Это достигается за счет подавления экспрессии CXCR4 на поверхности клеток-мишеней ВИЧ. Изоляты M-тропного ВИЧ-1, в которых используется только рецептор CCR5, называются R5; те, которые используют только CXCR4, называются X4, а те, которые используют оба, X4R5. Тем не менее, использование ко-рецепторов в одиночку не объясняет вирусный тропизм, так как не все вирусы R5 могут использовать CCR5 на макрофаги для продуктивной инфекции [45] и ВИЧ может также заразить подтип миелоидных дендритных клеток , [48] , которые вероятно, представляют собой резервуар , поддерживающий инфекцию, когда CD4+ Количество Т-клеток снизилось до чрезвычайно низкого уровня.
Некоторые люди устойчивы к определенным штаммам ВИЧ. [49] Например, люди с мутацией CCR5-Δ32 устойчивы к заражению вирусом R5, поскольку мутация не позволяет ВИЧ связываться с этим корецептором, что снижает его способность инфицировать клетки-мишени.
Половой акт - основной путь передачи ВИЧ. И X4, и R5 HIV присутствуют в семенной жидкости , что позволяет вирусу передаваться от мужчины его сексуальному партнеру . Затем вирионы могут инфицировать многочисленные клеточные мишени и распространяться по всему организму. Однако процесс отбора [ требуется дальнейшее объяснение ] приводит к преимущественной передаче вируса R5 по этому пути. [50] [51] [52] У пациентов, инфицированных ВИЧ-1 подтипа B, часто наблюдается переключение корецепторов на поздних стадиях заболевания и Т-тропные варианты, которые могут инфицировать различные Т-клетки через CXCR4. [53]Затем эти варианты реплицируются более агрессивно с повышенной вирулентностью, что вызывает быстрое истощение Т-клеток, коллапс иммунной системы и оппортунистические инфекции, знаменующие появление СПИДа. [54] ВИЧ-инфицированные пациенты заражаются чрезвычайно широким спектром оппортунистических инфекций, что было особенно проблематично до начала терапии ВААРТ ; тем не менее, те же самые инфекции регистрируются среди ВИЧ-инфицированных пациентов, обследованных после смерти после начала антиретровирусной терапии. [3]Таким образом, в ходе инфекции адаптация вируса к использованию CXCR4 вместо CCR5 может быть ключевым этапом в прогрессировании СПИДа. Ряд исследований с участием людей, инфицированных подтипом B, показал, что от 40 до 50 процентов пациентов со СПИДом могут быть носителями вирусов SI и, как предполагается, фенотипа X4. [55] [56]
ВИЧ-2 гораздо менее патогенен, чем ВИЧ-1, и его распространение во всем мире ограничено Западной Африкой . Принятие ВИЧ-2 «дополнительных генов» и его более беспорядочная модель использования корецепторов (включая независимость от CD4) может помочь вирусу в его адаптации, чтобы избежать врожденных факторов рестрикции, присутствующих в клетках-хозяевах. Адаптация к использованию нормальных клеточных механизмов для передачи и продуктивной инфекции также способствовала репликации ВИЧ-2 у людей. Стратегия выживания любого инфекционного агента состоит не в том, чтобы убить своего хозяина, а в конечном итоге стать комменсальным организмом. Достигнув низкой патогенности, со временем будут отобраны варианты, которые более успешны при передаче. [57]
Цикл репликации
Вход в камеру
Вирионов ВИЧ попадает макрофагов и CD4 + Т - клеток с помощью адсорбции из гликопротеинов на ее поверхности с рецепторами на клетке - мишени с последующим слиянием вирусной оболочки с целевой клеточной мембраной и высвобождения капсида ВИЧ в клетку. [58] [59]
Вход в клетку начинается посредством взаимодействия комплекса тримерной оболочки ( спайк gp160 ) с вирусной оболочкой ВИЧ и как CD4, так и корецептора хемокина (обычно CCR5 или CXCR4 , но, как известно, взаимодействуют другие) на поверхности клетки-мишени. [58] [59] Gp120 связывается с интегрином α 4 β 7, активируя LFA-1 , центральный интегрин, участвующий в создании вирусологических синапсов , которые способствуют эффективному распространению ВИЧ-1 от клетки к клетке. [60] Спайк gp160 содержит связывающие домены как для CD4, так и для хемокиновых рецепторов. [58][59]
Первый этап слияния включает высокоаффинное присоединение CD4-связывающих доменов gp120 к CD4. Как только gp120 связывается с белком CD4, комплекс оболочки претерпевает структурные изменения, обнажая связывающие домены хемокинового рецептора gp120 и позволяя им взаимодействовать с целевым хемокиновым рецептором. [58] [59] Это обеспечивает более стабильное двустороннее прикрепление, которое позволяет N-концевому слитому пептиду gp41 проникать через клеточную мембрану. [58] [59] Повторять последовательностив gp41, HR1 и HR2 затем взаимодействуют, вызывая коллапс внеклеточной части gp41 в форму шпильки. Эта петлевая структура сближает вирус и клеточные мембраны, обеспечивая слияние мембран и последующее проникновение вирусного капсида. [58] [59]
После связывания ВИЧ с клеткой-мишенью в клетку вводятся РНК ВИЧ и различные ферменты, включая обратную транскриптазу, интегразу, рибонуклеазу и протеазу. [58] [ неудавшаяся проверка ] Во время переноса микротрубочек в ядро вирусный геном однонитевой РНК транскрибируется в двухцепочечную ДНК, которая затем интегрируется в хромосому хозяина.
ВИЧ может инфицировать дендритные клетки (ДК) этим путем CD4-CCR5, но также может быть использован другой путь с использованием маннозо-специфических рецепторов лектина С-типа, таких как DC-SIGN . [61] ДК - одни из первых клеток, с которыми вирус сталкивается при передаче половым путем. В настоящее время считается, что они играют важную роль, передавая ВИЧ в Т-клетки, когда вирус захватывается слизистой оболочкой DC. [61] Считается , что присутствие FEZ-1 в нейронах предотвращает инфицирование клеток ВИЧ. [62]
Долгое время считалось, что проникновение ВИЧ-1, как и проникновение многих других ретровирусов, происходит исключительно через плазматическую мембрану. Однако совсем недавно сообщалось о продуктивной инфекции посредством pH- независимого, опосредованного клатрином эндоцитоза ВИЧ-1, и недавно было предложено, что это единственный путь продуктивного проникновения. [63] [64] [65] [66] [67]
Репликация и транскрипция
Вскоре после того, как вирусный капсид попадает в клетку, фермент, называемый обратной транскриптазой, освобождает геном одноцепочечной РНК с положительным смыслом от прикрепленных вирусных белков и копирует его в молекулу комплементарной ДНК (кДНК). [68] Процесс обратной транскрипции чрезвычайно подвержен ошибкам, и возникающие в результате мутации могут вызвать лекарственную устойчивость или позволить вирусу ускользнуть от иммунной системы организма. Обратная транскриптаза также обладает рибонуклеазной активностью, которая разрушает вирусную РНК во время синтеза кДНК, а также ДНК-зависимой активностью ДНК-полимеразы, которая создает смысловую ДНК из антисмысловой кДНК. [69]Вместе кДНК и ее комплемент образуют двухцепочечную вирусную ДНК, которая затем транспортируется в ядро клетки . Интеграция вирусной ДНК в геном клетки-хозяина осуществляется другим вирусным ферментом, называемым интегразой . [68]
Интегрированная вирусная ДНК может тогда находиться в спящем состоянии на латентной стадии ВИЧ-инфекции. [68] Для активного продуцирования вируса необходимо присутствие определенных клеточных факторов транскрипции , наиболее важным из которых является NF- κ B (ядерный фактор каппа B), который активируется при активации Т-клеток. [70] Это означает, что клетки, которые, скорее всего, станут мишенью, проникнуты и впоследствии убиты ВИЧ, - это клетки, активно борющиеся с инфекцией.
Во время вирусной репликации, интегрированной ДНК провируса является транскрибируется в РНК, некоторые из которых затем подвергаются сплайсинг РНК для получения зрелых матричными РНК (мРНК). Этот мРНК экспортируется из ядра в цитоплазму , где они переведенная в регуляторных белках Tat (что стимулирует новые производства вируса) и Rev . По мере того, как продуцируется только что продуцируемый белок Rev, он перемещается в ядро, где связывается с полноразмерными, несплицированными копиями вирусных РНК и позволяет им покинуть ядро. [71]Некоторые из этих полноразмерных РНК функционируют как новые копии вирусного генома, тогда как другие функционируют как мРНК, которые транслируются с образованием структурных белков Gag и Env. Белки Gag связываются с копиями генома вирусной РНК, упаковывая их в новые вирусные частицы. [72]
ВИЧ-1 и ВИЧ-2, по-видимому, по-разному упаковывают свою РНК. [73] [74] ВИЧ-1 будет связываться с любой подходящей РНК. [75] ВИЧ-2 будет предпочтительно связываться с мРНК, которая использовалась для создания самого белка Gag. [76]
Рекомбинация
Два генома РНК заключены в каждую частицу ВИЧ-1 (см. Структура и геном ВИЧ ). При инфицировании и репликации, катализируемых обратной транскриптазой, может происходить рекомбинация между двумя геномами. [77] [78] Рекомбинация происходит, когда однонитевые геномы с положительной РНК подвергаются обратной транскрипции с образованием ДНК. Во время обратной транскрипции формирующаяся ДНК может многократно переключаться между двумя копиями вирусной РНК. Эта форма рекомбинации известна как выбор копии. События рекомбинации могут происходить по всему геному. В каждом цикле репликации может происходить от двух до 20 событий рекомбинации на геном, и эти события могут быстро перетасовать генетическую информацию, которая передается от родительских геномов к потомкам. [78]
Рекомбинация вирусов вызывает генетические вариации, которые, вероятно, вносят вклад в развитие устойчивости к антиретровирусной терапии . [79] Рекомбинация также может в принципе способствовать преодолению иммунной защиты хозяина. Тем не менее, для реализации адаптивных преимуществ генетической изменчивости, два вирусных генома, упакованные в отдельные вирусные частицы, должны возникать из отдельных родительских родительских вирусов с разной генетической структурой. Неизвестно, как часто такая смешанная упаковка встречается в естественных условиях. [80]
Bonhoeffer et al. [81] предположили, что переключение матрицы с помощью обратной транскриптазы действует как процесс репарации для устранения разрывов в геноме одноцепочечной РНК. Кроме того, Ху и Темин [77]предположили, что рекомбинация является адаптацией для восстановления повреждений в геномах РНК. Переключение цепи (рекомбинация с выбором копии) с помощью обратной транскриптазы может генерировать неповрежденную копию геномной ДНК из двух поврежденных копий генома одноцепочечной РНК. Такой взгляд на адаптивное преимущество рекомбинации при ВИЧ может объяснить, почему каждая частица ВИЧ содержит два полных генома, а не один. Более того, точка зрения, что рекомбинация - это процесс репарации, подразумевает, что польза от репарации может проявляться в каждом цикле репликации, и что эта польза может быть реализована независимо от того, различаются ли два генома генетически. С точки зрения того, что рекомбинация при ВИЧ является процессом репарации, генерация рекомбинационной вариации будет следствием, но не причиной эволюции переключения матрицы. [81]
Инфекция ВИЧ-1 вызывает хроническое воспаление и выработку активных форм кислорода . [82] Таким образом, геном ВИЧ может быть уязвим для окислительных повреждений , включая разрывы одноцепочечной РНК. Для ВИЧ, как и для вирусов в целом, успешное инфицирование зависит от преодоления защитных стратегий хозяина, которые часто включают производство активных форм кислорода, повреждающих геном. Таким образом, Michod et al. [83] предположили, что рекомбинация с помощью вирусов - это адаптация для восстановления повреждений генома, а рекомбинационная вариация - это побочный продукт, который может дать отдельную пользу.
Сборка и выпуск
Заключительный этап вирусного цикла - сборка новых вирионов ВИЧ-1 - начинается на плазматической мембране клетки-хозяина. Полипротеина Env (gp160) проходит через эндоплазматической сети и транспортируется в аппарат Гольджи , где он расщепляется с помощью фурин в результате чего два гликопротеины оболочки ВИЧ, gp41 и gp120 . [84]Они транспортируются к плазматической мембране клетки-хозяина, где gp41 закрепляет gp120 на мембране инфицированной клетки. Полипротеины Gag (p55) и Gag-Pol (p160) также связываются с внутренней поверхностью плазматической мембраны вместе с геномной РНК ВИЧ, когда формирующийся вирион начинает отрастать от клетки-хозяина. Почковавший вирион все еще незрелый, так как полипротеины gag все еще должны быть расщеплены на фактические белки матрицы, капсида и нуклеокапсида. Это расщепление опосредуется упакованной вирусной протеазой и может быть ингибировано антиретровирусными препаратами класса ингибиторов протеаз . Затем различные структурные компоненты собираются, чтобы произвести зрелый вирион ВИЧ. [85] Только зрелые вирионы могут инфицировать другую клетку.
Распространение по телу
Классический процесс заражения клетки вирионом можно назвать «бесклеточным распространением», чтобы отличить его от недавно признанного процесса, называемого «межклеточное распространение». [86] При внеклеточном распространении (см. Рисунок) вирусные частицы зарождаются из инфицированной Т-клетки, попадают в кровь или внеклеточную жидкость, а затем заражают другую Т-клетку после случайного контакта. [86] ВИЧ может также распространяться путем прямой передачи от одной клетки к другой в процессе распространения от клетки к клетке, для чего были описаны два пути. Во-первых, инфицированная Т-клетка может передавать вирус непосредственно целевой Т-клетке через вирусологический синапс . [60] [87] Во-вторых, антигенпрезентирующая клетка (APC)такие как макрофаг или дендритная клетка, могут передавать ВИЧ к Т-клеткам посредством процесса, который включает либо продуктивную инфекцию (в случае макрофагов), либо захват и перенос вирионов в транс (в случае дендритных клеток). [88] Какой бы путь ни использовался, сообщается, что инфицирование путем передачи вируса от клетки к клетке намного эффективнее, чем распространение вируса вне клетки. [89] Этому повышению эффективности способствует ряд факторов, в том числе отпочкование поляризованного вируса к месту межклеточного контакта, близкое расположение клеток, которое минимизирует диффузию вирионов в жидкой фазе , и кластеризацию рецепторов входа ВИЧ на целевая ячейка в сторону зоны контакта. [87]Считается, что распространение от клетки к клетке особенно важно в лимфоидных тканях, где CD4 + Т-клетки плотно упакованы и могут часто взаимодействовать. [86] Исследования прижизненной визуализации подтвердили концепцию вирусологического синапса ВИЧ in vivo . [90] Многие механизмы распространения ВИЧ способствуют продолжающейся репликации вируса, несмотря на антиретровирусную терапию. [86] [91]
Генетическая изменчивость
ВИЧ отличается от многих вирусов очень высокой генетической изменчивостью . Это разнообразие является результатом его быстрого цикла репликации , с генерацией около 10 10 вирионов каждый день, в сочетании с высокой скоростью мутаций примерно 3 x 10 -5 на нуклеотидное основание на цикл репликации и рекомбиногенными свойствами обратной транскриптазы. [92] [93] [94]
Этот сложный сценарий приводит к появлению множества вариантов ВИЧ у одного инфицированного пациента в течение одного дня. [92] Эта вариабельность усугубляется, когда одна клетка одновременно заражается двумя или более разными штаммами ВИЧ. При одновременном заражении геном вирионов потомства может состоять из цепей РНК двух разных штаммов. Затем этот гибридный вирион заражает новую клетку, где подвергается репликации. Когда это происходит, обратная транскриптаза, перепрыгивая между двумя различными матрицами РНК, будет генерировать вновь синтезированную последовательность ретровирусной ДНК, которая является рекомбинантной между двумя родительскими геномами. [92] Эта рекомбинация наиболее очевидна, когда она происходит между подтипами.[92]
Тесно связаны вирус иммунодефицита обезьян (ВИО) превратилась в много штаммов, классифицированных по природным видов хозяев. Считается, что штаммы SIV африканской зеленой обезьяны (SIVagm) и сажистого мангабея (SIVsmm) имеют долгую историю эволюции со своими хозяевами. Эти хозяева адаптировались к присутствию вируса [95], который присутствует в больших количествах в крови хозяина, но вызывает только умеренный иммунный ответ [96] , не вызывает развития обезьяньего СПИДа [97] и вызывает не подвергаться обширным мутациям и рекомбинации, типичным для ВИЧ-инфекции у людей. [98]
Напротив, когда эти штаммы заражают виды, которые не адаптировались к SIV («гетерологичные» или похожие хозяева, такие как макаки-резусы или циномологи ), у животных развивается СПИД, и вирус генерирует генетическое разнообразие, подобное тому, которое наблюдается при ВИЧ-инфекции человека. [99] Шимпанзе SIV (SIVcpz), ближайший генетический родственник ВИЧ-1, связан с повышенной смертностью и симптомами, подобными СПИДу, у его естественного хозяина. [100] SIVcpz, по-видимому, относительно недавно был передан популяциям шимпанзе и людей, поэтому их хозяева еще не адаптировались к вирусу. [95] Этот вирус также потерял функцию nefген, который присутствует в большинстве SIV. Для непатогенных вариантов SIV nef подавляет активацию Т-клеток через маркер CD3 . Функция Nef в непатогенных формах SIV заключается в подавлении экспрессии воспалительных цитокинов , MHC-1 и сигналов, влияющих на транспорт Т-клеток. В ВИЧ-1 и SIVcpz nef не ингибирует активацию Т-клеток и утратил эту функцию. Без этой функции более вероятно истощение Т-лимфоцитов, что приведет к иммунодефициту. [100] [101]
На основе различий в области оболочки ( env ) были идентифицированы три группы ВИЧ-1 : M, N и O. [102] Группа M является наиболее распространенной и подразделяется на восемь подтипов (или кладов ) на основе на весь геном, которые различаются географически. [103] Наиболее распространены подтипы B (в основном в Северной Америке и Европе), A и D (в основном в Африке) и C (в основном в Африке и Азии); эти подтипы образуют ветви в филогенетическом дереве, представляющем происхождение группы M ВИЧ-1. Коинфекцияс различными подтипами дает начало циркулирующим рекомбинантным формам (CRF). В 2000 году, последнем году, в котором был проведен анализ глобальной распространенности подтипа, 47,2% инфекций во всем мире относились к подтипу C, 26,7% - к подтипу A / CRF02_AG, 12,3% - к подтипу B, 5,3% - к подтипу D, 3,2% относились к CRF_AE, а остальные 5,3% составляли другие подтипы и CRF. [104] Большинство исследований ВИЧ-1 сосредоточено на подтипе B; несколько лабораторий специализируются на других подтипах. [105] Существование четвертой группы, «P», было выдвинуто на основании вируса, изолированного в 2009 году. [106] Штамм, по-видимому, произошел от гориллы SIV (SIVgor), впервые выделенной из западных низинных горилл в 2006 году [ 106] . 106]
Ближайшим родственником ВИЧ-2 является SIVsm, штамм SIV, обнаруженный в сажистых мангабах. Поскольку ВИЧ-1 происходит от SIVcpz, а ВИЧ-2 - от SIVsm, генетическая последовательность ВИЧ-2 лишь частично гомологична ВИЧ-1 и больше напоминает последовательность SIVsm. [ необходима цитата ] [107]
Диагностика
Многие ВИЧ-положительные люди не знают, что они инфицированы вирусом. [108] Например, в 2001 году менее 1% сексуально активного городского населения в Африке прошли тестирование, а в сельской местности эта доля еще ниже. [108] Кроме того, в 2001 году только 0,5% беременных женщин, посещающих городские медицинские учреждения, прошли консультации, прошли тестирование или получили результаты тестов. [108] Опять же, эта доля еще ниже в сельских медицинских учреждениях. [108] Поскольку доноры могут не знать о своей инфекции, донорская кровь и продукты крови, используемые в медицине и медицинских исследованиях , обычно проходят скрининг на ВИЧ. [109]
Первоначально тестирование на ВИЧ-1 проводится с использованием иммуноферментного анализа (ИФА) для выявления антител к ВИЧ-1. Образцы с нереактивным результатом первоначального ИФА считаются ВИЧ-отрицательными, если только не произошел новый контакт с инфицированным партнером или партнером с неизвестным ВИЧ-статусом. Образцы с реактивным результатом ELISA повторно тестируются в двух экземплярах. [110] Если результат любого повторного теста является реактивным, образец регистрируется как повторно реактивный и подвергается подтверждающему тестированию с помощью более специфического дополнительного теста (например, полимеразной цепной реакции (ПЦР), вестерн-блоттинга или, реже, иммунофлуоресценции. проба(ЕСЛИ)). Только образцы, которые повторно реактивны по данным ELISA и положительны по данным IFA или ПЦР или реактивны по данным вестерн-блоттинга, считаются ВИЧ-положительными и указывают на ВИЧ-инфекцию. Образцы, которые повторно реагируют на ELISA, иногда дают неопределенный результат вестерн-блоттинга, который может быть либо неполным ответом антител на ВИЧ у инфицированного человека, либо неспецифическими реакциями у неинфицированного человека. [111]
Смертность от ВИЧ (кроме США) в 2014 г. [112]
Хотя ИФА можно использовать для подтверждения инфекции в этих неоднозначных случаях, этот анализ не получил широкого распространения. Как правило, второй образец следует собирать более чем через месяц и повторно тестировать для лиц с неопределенными результатами вестерн-блоттинга. Тестирование нуклеиновых кислот (например, вирусная РНК или метод амплификации провирусной ДНК), хотя и менее широко доступно, также может помочь в диагностике в определенных ситуациях. [110] Кроме того, несколько протестированных образцов могут дать неубедительные результаты из-за малого количества образцов. В этих ситуациях берется второй образец и тестируется на ВИЧ-инфекцию.
Современное тестирование на ВИЧ чрезвычайно точное, если принять во внимание период окна . Один скрининговый тест является правильным более чем в 99% случаев. [113] Вероятность ложноположительного результата при стандартном двухэтапном протоколе тестирования оценивается примерно в 1 на 250 000 в популяции с низким уровнем риска. [114] Тестирование после контакта рекомендуется немедленно, а затем через шесть недель, три месяца и шесть месяцев. [115]
Последние рекомендации Центров по контролю и профилактике заболеваний США (CDC) показывают, что тестирование на ВИЧ должно начинаться с комбинированного иммуноанализа на антитела к ВИЧ-1 и ВИЧ-2 и антиген p24 . Отрицательный результат исключает контакт с ВИЧ, в то время как положительный результат должен сопровождаться иммуноанализом на дифференцировку антител к ВИЧ-1/2, чтобы определить, какие антитела присутствуют. Это дает четыре возможных сценария:
- 1. ВИЧ-1 (+) и ВИЧ-2 (-): обнаружены антитела к ВИЧ-1.
- 2. ВИЧ-1 (-) и ВИЧ-2 (+): обнаружены антитела к ВИЧ-2.
- 3. ВИЧ-1 (+) и ВИЧ-2 (+): обнаружены антитела к ВИЧ-1 и ВИЧ-2.
- 4. ВИЧ-1 (-) или неопределенный и ВИЧ-2 (-): необходимо провести тест на нуклеиновую кислоту для выявления острой инфекции ВИЧ-1 или ее отсутствия. [116]
Исследование
Исследования ВИЧ / СПИДа включают в себя все медицинские исследования , направленные на предотвращение, лечение или излечение ВИЧ / СПИДа , а также фундаментальные исследования природы ВИЧ как инфекционного агента и СПИДа как заболевания, вызываемого ВИЧ.
Многие правительства и исследовательские учреждения участвуют в исследованиях по ВИЧ / СПИДу. Это исследование включает в себя вмешательства в области психического здоровья , такие как исследования в области полового воспитания и разработки лекарств , такие как исследования микробицидов для болезней, передаваемых половым путем , вакцин против ВИЧ и антиретровирусных препаратов . [117] Другие области медицинских исследований включают темы доконтактной профилактики , постконтактной профилактики , обрезания и ВИЧ , а также ускоренного старения .
Лечение и передача
Лечение ВИЧ / СПИДа обычно включает использование нескольких антиретровирусных препаратов . Во многих частях мира ВИЧ стал хроническим заболеванием, при котором прогрессирование до СПИДа становится все более редким.
Латентный период ВИЧ и, как следствие, резервуар вируса в CD4 + Т-клетках, дендритных клетках, а также макрофагах, является основным барьером на пути к уничтожению вируса. [19]
Важно отметить, что, хотя ВИЧ очень вирулентен, передача не происходит половым путем, если у ВИЧ-инфицированного человека неизменно неопределяемая вирусная нагрузка (<50 копий / мл) из-за антиретровирусного лечения. Впервые это было высказано Федеральной комиссией Швейцарии по СПИДу / ВИЧ в 2008 году в Заявлении Швейцарии , хотя в то время это заявление было спорным. [118] [119] Однако после многочисленных исследований стало ясно, что вероятность передачи ВИЧ половым путем фактически равна нулю, если у ВИЧ-инфицированного человека неизменно неопределяемая вирусная нагрузка; это известно как U = U, «Undetectable = Untransmittable», также выражается как «не могу передать это». [120] [121]Исследования, демонстрирующие U = U: Противоположности притягиваются, [122] ПАРТНЕР 1, [123] ПАРТНЕР 2, [5] [124] (для пар мужчина-мужчина) [125] и HPTN052 [126] (для гетеросексуальных пар), когда «Партнер, живущий с ВИЧ, имел длительно подавленную вирусную нагрузку». [125] В этих исследованиях участвовали пары, в которых один партнер был ВИЧ-положительным, а другой - ВИЧ-отрицательным, и проходили регулярные тесты на ВИЧ. Всего по четырем исследованиям было зарегистрировано 4097 пар на четырех континентах, и было зарегистрировано 151 880 половых актов без презерватива; не было ни одной филогенетически связанной передачи ВИЧ, когда у положительного партнера вирусная нагрузка не определялась. [127]После этого заявление о консенсусе U = U, поддерживающее использование «нулевого риска», было подписано сотнями людей и организаций, включая Центр контроля заболеваний США , Британскую ассоциацию по ВИЧ и медицинский журнал The Lancet . [128] Важность окончательных результатов исследования PARTNER 2 была описана медицинским директором Terrence Higgins Trust как «невозможно переоценить», а ведущий автор Элисон Роджер заявила, что сообщение о том, что «неопределяемая вирусная нагрузка делает ВИЧ непередаваемым». .. может помочь остановить пандемию ВИЧ, предотвратив передачу ВИЧ. [129] Авторы резюмировали свои выводы в The Lancet следующим образом: [5]
Наши результаты обеспечивают уровень доказательств подавления вируса и риска передачи ВИЧ для геев, аналогичный тому, который ранее был получен для гетеросексуальных пар, и предполагают, что риск передачи ВИЧ в однополых парах через секс без презерватива при подавлении вирусной нагрузки ВИЧ фактически равен нулю. Наши результаты подтверждают идею кампании U = U (необнаружимое равно непередаваемо) и преимущества раннего тестирования и лечения ВИЧ. [5]
Этот результат согласуется с выводом , представленный Энтони С. Фаучи , директор Национального института аллергии и инфекционных заболеваний для американских Национальных институтов здравоохранения , и его команда в точки зрения , опубликованной в журнале Американской медицинской ассоциации , что U = U - эффективный метод профилактики ВИЧ при сохранении неопределяемой вирусной нагрузки. [6] [125]
Реактивация генитального герпеса (HSV-2) у инфицированных этим вирусом приводит к увеличению количества CD4 + T-клеток, обогащенных CCR-5, а также воспалительных дендритных клеток в подслизистой основе кожи гениталий. Тропизм ВИЧ к CCR-5-позитивным клеткам объясняет двух-трехкратное увеличение заражения ВИЧ среди людей с генитальным герпесом. Ежедневный прием противовирусных препаратов (например, ацикловира) не уменьшает субклиническое постреактивационное воспаление и, следовательно, не снижает риск заражения ВИЧ. [130] [131]
История
Открытие
Первая новость о «новой экзотической болезни» появилась 18 мая 1981 года в газете для геев New York Native . [132]
СПИД был впервые клинически обнаружен в 1981 году в США. [133] Первоначальные случаи были группой потребителей инъекционных наркотиков и геев без известной причины нарушения иммунитета, у которых проявлялись симптомы пневмоцистной пневмонии (PCP или PJP, последний термин означает, что возбудитель теперь называется Pneumocystis jirovecii ), редкая оппортунистическая инфекция, которая, как известно, возникает у людей с очень ослабленной иммунной системой. [134] Вскоре после этого исследователи Медицинской школы Нью-Йоркского университета изучили мужчин-геев, у которых развился ранее редкий рак кожи, называемый саркомой Капоши (СК). [135] [136] Возникло еще много случаев PJP и KS, что стало предупреждением для США.Центры по контролю и профилактике заболеваний (CDC) и целевая группа CDC были сформированы для мониторинга вспышки. [137] Считается, что самый ранний ретроспективно описанный случай СПИДа произошел в Норвегии начиная с 1966 года. [138]
Вначале CDC не имел официального названия болезни, часто ссылаясь на нее посредством заболеваний, которые были с ней связаны, например, лимфаденопатии , болезни, в честь которой первооткрыватели ВИЧ первоначально назвали вирус. [139] [140] Они также использовали саркому Капоши и оппортунистические инфекции , название, под которым в 1981 году была создана целевая группа. [141] В общей прессе термин GRID , который обозначал иммунодефицит , связанный с геями , был придуман. [142] CDC, в поисках названия и изучая зараженные сообщества, придумал «болезнь 4H», как будто выделив гомосексуалистов, потребителей героина,гемофилики и гаитяне . [143] [144] Однако, после определения того, что СПИД не был изолирован для гей-сообщества , [141] стало понятно, что термин GRID вводит в заблуждение, и СПИД был представлен на встрече в июле 1982 года. [145] К сентябрю 1982 года CDC начал использовать название AIDS. [146]
В 1983 году две отдельные исследовательские группы во главе с американцем Робертом Галло и французскими исследователями Франсуазой Барре-Синусси и Люком Монтанье независимо друг от друга заявили, что новый ретровирус мог инфицировать больных СПИДом, и опубликовали свои выводы в том же номере журнала Science . [147] [148] [149] Галло утверждал, что вирус, который его группа выделила от человека со СПИДом, поразительно похож по форме на другие человеческие Т-лимфотропные вирусы.(HTLV) его группа была изолирована первой. В 1987 году Галло признал, что вирус, который, как он утверждал, обнаружил в 1984 году, на самом деле был вирусом, присланным ему из Франции годом ранее. [150] Группа Галло назвала свой недавно выделенный вирус HTLV-III. Группа Монтанье выделила вирус у пациента с отеком лимфатических узлов шеи и физической слабостью , двумя классическими симптомами первичной ВИЧ-инфекции. Вопреки отчету группы Галло, Монтанье и его коллеги показали, что коровые белки этого вируса иммунологически отличаются от белков HTLV-I. Группа Монтанье назвала свой изолированный вирус вирусом, связанным с лимфаденопатией (LAV). [137]Поскольку эти два вируса оказались одинаковыми, в 1986 году LAV и HTLV-III были переименованы в ВИЧ. [151]
Другой группой, работавшей одновременно с группами Монтанье и Галло, была группа доктора Джея Леви из Калифорнийского университета в Сан-Франциско . Он независимо открыл вирус СПИДа в 1983 году и назвал его СПИД-ассоциированным ретровирусом (АРВ). [152] Этот вирус сильно отличался от вируса, о котором сообщили группы Монтанье и Галло. Штаммы АРВ-препаратов впервые показали гетерогенность изолятов ВИЧ, и некоторые из них остаются классическими примерами вируса СПИДа, обнаруженного в Соединенных Штатах. [153]
Происхождение
Считается, что и ВИЧ-1, и ВИЧ-2 произошли от нечеловеческих приматов в Западной и Центральной Африке и, как полагают, передались человеку (процесс, известный как зооноз ) в начале 20 века. [154] [155]
ВИЧ-1, по-видимому, возник в южном Камеруне в результате эволюции SIVcpz, вируса иммунодефицита обезьян (SIV), который поражает диких шимпанзе (ВИЧ-1 происходит от эндемичного вируса SIVcpz у подвида шимпанзе Pan troglodytes troglodytes ). [156] [157] Ближайшим родственником ВИЧ-2 является SIVsmm, вирус сажистого мангабея ( Cercocebus atys atys ), обезьяны Старого Света, обитающей в прибрежной зоне Западной Африки (от южной части Сенегала до западной части Кот-д'Ивуара ). [21] Обезьяны Нового Света, такие как обезьяна-соваустойчивы к инфекции ВИЧ-1, возможно, из-за слияния двух генов вирусной устойчивости. [158]
Считается, что ВИЧ-1 преодолевал видовой барьер по крайней мере в трех отдельных случаях, давая начало трем группам вируса: M, N и O. [159]
Есть свидетельства того, что люди, которые участвуют в производстве мяса диких животных в качестве охотников или продавцов мяса диких животных, обычно приобретают SIV. [160] Однако SIV - слабый вирус, и иммунная система человека обычно подавляет его в течение нескольких недель после заражения. Считается, что необходимо несколько последовательных передач вируса от человека к человеку, чтобы дать ему достаточно времени для мутации в ВИЧ. [161] Кроме того, из-за относительно низкой скорости передачи от человека к человеку, он может распространяться среди населения только при наличии одного или нескольких каналов передачи высокого риска, которые, как считается, отсутствовали в Африке до 20 век.
Конкретные предлагаемые каналы передачи высокого риска, позволяющие вирусу адаптироваться к людям и распространяться в обществе, зависят от предполагаемого времени перехода от животного к человеку. Генетические исследования вируса предполагают, что самый недавний общий предок группы ВИЧ-1 M датируется примерно 1910 годом. [162] Сторонники этой датировки связывают эпидемию ВИЧ с возникновением колониализма и ростом крупных колониальных африканских городов, ведущими к социальным изменениям, включая различные модели половых контактов (особенно множественные, одновременные партнерства), распространение проституции и сопутствующую высокую частоту язвенных заболеваний половых органов (таких как сифилис ) в формирующихся колониальных городах. [163]Хотя скорость передачи ВИЧ при вагинальном половом акте обычно невысока, она многократно увеличивается, если один из партнеров страдает инфекцией, передающейся половым путем, что приводит к язвам на гениталиях. Колониальные города начала 1900-х годов были известны своей высокой распространенностью проституции и генитальных язв до такой степени, что с 1928 года считалось, что до 45% жителей восточного Леопольдвиля были проститутками, а с 1933 года - около 15% всех жителей города. этот же город был заражен одной из форм сифилиса . [163]
Самый ранний, хорошо задокументированный случай ВИЧ-инфекции у человека относится к 1959 году в Бельгийском Конго . [164] Вирус мог присутствовать в Соединенных Штатах еще в середине-конце 1950-х годов, когда шестнадцатилетний мужчина проявил симптомы в 1966 году и умер в 1969 году. [165]
Альтернативная точка зрения - не подкрепленная доказательствами - утверждает, что небезопасная медицинская практика в Африке в годы после Второй мировой войны, такая как нестерильное повторное использование одноразовых шприцев во время массовых кампаний вакцинации, антибиотиков и лечения малярии, была первоначальным вектором, позволившим вирус адаптироваться к людям и распространяться. [161] [166] [167]
Смотрите также
- Противовирусный препарат
- Открытие и разработка ингибиторов протеазы ВИЧ
- Отрицание ВИЧ / СПИДа
- Всемирный день борьбы со СПИДом
Рекомендации
- Перейти ↑ Weiss RA (май 1993 г.). «Как ВИЧ вызывает СПИД?». Наука . 260 (5112): 1273–9. Bibcode : 1993Sci ... 260.1273W . DOI : 10.1126 / science.8493571 . PMID 8493571 .
- ^ Douek DC, Редерер M, Koup RA (2009). «Новые концепции иммунопатогенеза СПИДа» . Ежегодный обзор медицины . 60 : 471–84. DOI : 10.1146 / annurev.med.60.041807.123549 . PMC 2716400 . PMID 18947296 .
- ^ a b Пауэлл М.К., Бенкова К., Селинджер П., Догоши М., Кинкорова Лунячкова I, Кутникова Х, Лаштикова Ю., Рубичкова А., Шпуркова З, Лаклова Л., Эйс В., Шах Дж, Хенеберг П. (2016). «Оппортунистические инфекции у ВИЧ-инфицированных пациентов сильно различаются по частотам и спектрам между пациентами с низким содержанием CD4 + клеток, обследованными после смерти, и пациентами, подвергнутыми компенсации, обследованными до смерти, независимо от эры ВААРТ» . PLOS ONE . 11 (9): e0162704. Bibcode : 2016PLoSO..1162704P . DOI : 10.1371 / journal.pone.0162704 . PMC 5017746 . PMID 27611681 .
- ^ ЮНЭЙДС , ВОЗ (декабрь 2007 г.). «Обновленная информация об эпидемии СПИДа 2007 г.» (PDF) . п. 10. Архивировано из оригинального (PDF) 22 ноября 2008 года . Проверено 12 марта 2008 года .
- ^ a b c d Роджер, Элисон Дж .; Камбиано, Валентина; Бруун, Тина; Вернацца, Пьетро; Коллинз, Саймон; Деген, Олаф; и другие. (2019). «Риск передачи ВИЧ через секс без презерватива в серодифференцированных гомосексуальных парах с ВИЧ-положительным партнером, получающим супрессивную антиретровирусную терапию (ПАРТНЕР): окончательные результаты многоцентрового проспективного обсервационного исследования» . Ланцет . 393 (10189): 2428–2438. DOI : 10.1016 / S0140-6736 (19) 30418-0 . PMC 6584382 . PMID 31056293 .
- ^ a b Eisinger, Роберт В .; Dieffenbach, Carl W .; Фаучи, Энтони С. (2019). «Вирусная нагрузка ВИЧ и трансмиссивность ВИЧ-инфекции: неопределяемое равно непередаваемое». JAMA . 321 (5): 451–452. DOI : 10,1001 / jama.2018.21167 . PMID 30629090 .
- ^ Mabuka Дж, Nduati R, Одем-Дэвис К, Д Петерсон, Овербо J (2012). Desrosiers RC (ред.). «ВИЧ-специфические антитела, способные вызывать ADCC, распространены в грудном молоке и связаны с пониженным риском передачи инфекции у женщин с высокой вирусной нагрузкой» . PLOS Патогены . 8 (6): e1002739. DOI : 10.1371 / journal.ppat.1002739 . PMC 3375288 . PMID 22719248 .
- ^ Хан, Роберт А .; Инхорн, Марсия Клэр, ред. (2009). Антропология и общественное здоровье: преодоление различий в культуре и обществе (2-е изд.). Оксфорд: Издательство Оксфордского университета. п. 449. ISBN. 978-0-19-537464-3. OCLC 192042314 .
- Перейти ↑ Mead MN (2008). «Загрязняющие вещества в грудном молоке: сравнение рисков и преимуществ грудного вскармливания» . Перспективы гигиены окружающей среды . 116 (10): A426–34. DOI : 10.1289 / ehp.116-A426 . PMC 2569122 . PMID 18941560 . Архивировано из оригинала 6 ноября 2008 года.
- ^ «Предотвращение передачи ВИЧ от матери ребенку» . HIV.gov . 15 мая 2017 года . Проверено 8 декабря 2017 года .
 Эта статья включает текст из этого источника, который находится в общественном достоянии .
Эта статья включает текст из этого источника, который находится в общественном достоянии . - Перейти ↑ Cunningham AL, Donaghy H, Harman AN, Kim M, Turville SG (2010). «Манипулирование вирусами функцией дендритных клеток». Текущее мнение в микробиологии . 13 (4): 524–529. DOI : 10.1016 / j.mib.2010.06.002 . PMID 20598938 .
- ^ Дойтш, Гилад; Галлоуэй, Николь ЛК; Гэн, Синь; Ян, Чжиюань; Монро, Кэтрин М .; Зепеда, Орландо; Хант, Питер В .; Хатано, Хирою; Совински, Стефани; Муньос-Ариас, Иса; Грин, Уорнер С. (2014). «Гибель клеток в результате пироптоза приводит к истощению Т-лимфоцитов CD4 при ВИЧ-1 инфекции» . Природа . 505 (7484): 509–514. Bibcode : 2014Natur.505..509D . DOI : 10,1038 / природа12940 . PMC 4047036 . PMID 24356306 .
- ^ Garg Н, Моль J, Джоши А (9 ноября 2012). «ВИЧ-1-индуцированный апоптоз свидетеля» . Вирусы . 4 (11): 3020–43. DOI : 10,3390 / v4113020 . PMC 3509682 . PMID 23202514 .
- ^ Кумар, Вини (2012). Базовая патология Роббинса (9-е изд.). п. 147. ISBN. 978-1-4557-3787-1.
- ^ Международный комитет по таксономии вирусов (2002). «61.0.6. Лентивирус» . Национальные институты здоровья . Проверено 28 февраля 2006 года .
- ^ Международный комитет по таксономии вирусов (2002). «61. Retroviridae» . Национальные институты здоровья . Проверено 28 февраля 2006 года .
- ^ Леви JA (1993). «Патогенез ВИЧ и долгосрочное выживание». СПИД . 7 (11): 1401–10. DOI : 10.1097 / 00002030-199311000-00001 . PMID 8280406 .
- Перейти ↑ Smith JA, Daniel R (2006). «По пути вируса: использование ретровирусами механизмов репарации ДНК хозяина». ACS Химическая биология . 1 (4): 217–26. DOI : 10.1021 / cb600131q . PMID 17163676 .
- ^ a b Силичиано, РФ; Грин, WC (2011). «Латентность ВИЧ» . Перспективы Колд-Спринг-Харбор в медицине . 1 (1): a007096. DOI : 10.1101 / cshperspect.a007096 . PMC 3234450 . PMID 22229121 .
- ^ Gilbert PB, McKeague IW, Эйзен G, Mullins C, Гуйе-НДИАЙЕ A, S Mboup, Kanki PJ (28 февраля 2003). «Сравнение инфекционности ВИЧ-1 и ВИЧ-2 из проспективного когортного исследования в Сенегале». Статистика в медицине . 22 (4): 573–593. DOI : 10.1002 / sim.1342 . PMID 12590415 .
- ^ a b Ривз Дж. Д., Домс Р. В. (2002). «Вирус иммунодефицита человека 2 типа» . Журнал общей вирусологии . 83 (Pt 6): 1253–65. DOI : 10.1099 / 0022-1317-83-6-1253 . PMID 12029140 .
- ^ Макговерн SL, Caselli E, N Григорьев, Shoichet BK (2002). «Общий механизм, лежащий в основе беспорядочных связей с ингибиторами виртуального и высокопроизводительного скрининга». Журнал медицинской химии . 45 (8): 1712–22. DOI : 10.1021 / jm010533y . hdl : 11380/977912 . PMID 11931626 .
- ^ По сравнению с обзором в: Фишер, Брюс; Харви, Ричард П .; Чампе, Памела С. (2007). Иллюстрированные обзоры Липпинкотта: микробиология . Иллюстрированные обзоры Липпинкотта. Хагерстаун, Мэриленд: Липпинкотт Уильямс и Уилкинс. п. 3. ISBN 978-0-7817-8215-9.
- ^ Б с д е е г Различные (2008). Сборник последовательностей ВИЧ, 2008 г. Введение (PDF) . Проверено 31 марта 2009 года .
- ^ a b c Чан, округ Колумбия, Фасс Д., Бергер Дж. М., Ким П. С. (1997). «Основная структура gp41 из гликопротеина оболочки ВИЧ» (PDF) . Cell . 89 (2): 263–73. DOI : 10.1016 / S0092-8674 (00) 80205-6 . PMID 9108481 . S2CID 4518241 .
- ^ Кляйн, Джошуа С .; Bjorkman, Pamela J .; Ралл, Гленн Ф. (27 мая 2010 г.). «Немного и далеко между: как ВИЧ может избежать авидности антител» . PLOS Патогены . 6 (5): e1000908. DOI : 10.1371 / journal.ppat.1000908 . PMC 2877745 . PMID 20523901 .
- ↑ Национальный институт здоровья (17 июня 1998 г.). «Кристаллическая структура ключевого белка ВИЧ открывает новые возможности для профилактики и лечения» (пресс-релиз). Архивировано из оригинального 19 -го февраля 2006 года . Проверено 14 сентября 2006 года .
- ↑ Беренс, Анна-Янина; Васильевич, Снежана; Причард, Лаура К.; Харви, Дэвид Дж; Андев, Раджиндер С; Крумм, Стефани А; Струве, Уэстон Б; Купо, Альберт; Кумар, Абхинав; Зицманн, Николь; Сибрайт, Джемма Э; Крамер, Хольгер Б; Спенсер, Дэниел И. Р.; Ройл, Луиза; Ли, Чон Хён; Klasse, Per J; Бертон, Деннис Р.; Уилсон, Ян А; Уорд, Эндрю Б; Сандерс, Роджер В; Мур, Джон П.; Дур, Кэти Дж; Криспин, Макс (2016). «Состав и антигенные эффекты отдельных гликановых сайтов тримерного гликопротеина оболочки ВИЧ-1» . Сотовые отчеты . 14 (11): 2695–706. DOI : 10.1016 / j.celrep.2016.02.058 . PMC 4805854 . PMID 26972002 .
- ^ Причард, Лаура К; Спенсер, Дэниел И. Р.; Ройл, Луиза; Бономелли, Камилла; Сибрайт, Джемма Э; Беренс, Анна-Янина; Кулп, Дэниел В; Менис, Сергей; Крумм, Стефани А; Данлоп, Д. Кэмерон; Криспин, Дэниел Дж; Боуден, Томас А; Сканлан, Кристофер Н; Уорд, Эндрю Б; Schief, William R; Дур, Кэти Дж; Криспин, Макс (2015). «Кластеризация гликанов стабилизирует маннозный участок ВИЧ-1 и сохраняет уязвимость для широко нейтрализующих антител» . Nature Communications . 6 : 7479. Bibcode : 2015NatCo ... 6.7479P . DOI : 10.1038 / ncomms8479 . PMC 4500839 . PMID 26105115 .
- ^ Причард, Лаура К; Харви, Дэвид Дж; Бономелли, Камилла; Криспин, Макс; Дур, Кэти Дж (2015). «Клеточно-белковое гликозилирование нативной расщепленной оболочки ВИЧ-1» . Журнал вирусологии . 89 (17): 8932–44. DOI : 10,1128 / JVI.01190-15 . PMC 4524065 . PMID 26085151 .
- ^ Криспин, Макс; Дур, Кэти Дж (2015). «Нацеливание гликанов хозяина на вирусы с оболочкой для создания вакцины на основе антител» . Текущее мнение в вирусологии . 11 : 63–9. DOI : 10.1016 / j.coviro.2015.02.002 . PMC 4827424 . PMID 25747313 .
- ^ Жюльен, Жан-Филипп; Купо, Альберт; Сок, Девин; Stanfield, Robyn L .; Люмкис, Дмитрий; Deller, Marc C .; Классе, Пер-Йохан; Бертон, Деннис Р .; Сандерс, Роджер В. (20 декабря 2013 г.). «Кристаллическая структура растворимого расщепленного тримера оболочки ВИЧ-1» . Наука . 342 (6165): 1477–1483. Bibcode : 2013Sci ... 342.1477J . DOI : 10.1126 / science.1245625 . ISSN 1095-9203 . PMC 3886632 . PMID 24179159 .
- ^ Lyumkis Дмитрий; Жюльен, Жан-Филипп; де Валь, Наталья; Купо, Альберт; Поттер, Клинтон С .; Классе, Пер-Йохан; Бертон, Деннис Р .; Сандерс, Роджер В .; Мур, Джон П. (20 декабря 2013 г.). «Крио-ЭМ структура полностью гликозилированного растворимого расщепленного тримера оболочки ВИЧ-1» . Наука . 342 (6165): 1484–1490. Bibcode : 2013Sci ... 342.1484L . DOI : 10.1126 / science.1245627 . ISSN 1095-9203 . PMC 3954647 . PMID 24179160 .
- ^ Сандерс, Роджер В .; Деркинг, Рональд; Купо, Альберт; Жюльен, Жан-Филипп; Ясмин, Анила; де Валь, Наталья; Ким, Хелен Дж .; Блаттнер, Клаудиа; де ла Пенья, Альба Торрентс (1 сентября 2013 г.). «Расщепленный растворимый тример Env ВИЧ-1 нового поколения, BG505 SOSIP.664 gp140, экспрессирует несколько эпитопов для широко нейтрализующих, но не ненейтрализующих антител» . PLOS Патогены . 9 (9): e1003618. DOI : 10.1371 / journal.ppat.1003618 . ISSN 1553-7374 . PMC 3777863 . PMID 24068931 .
- ^ Причард, Лаура К .; Васильевич, Снежана; Озоровский, Габриэль; Сибрайт, Джемма Э .; Купо, Альберт; Ринге, Раджеш; Ким, Хелен Дж .; Сандерс, Роджер В .; Дур, Кэти Дж. (16 июня 2015 г.). «Структурные ограничения определяют гликозилирование тримеров оболочки ВИЧ-1» . Сотовые отчеты . 11 (10): 1604–1613. DOI : 10.1016 / j.celrep.2015.05.017 . ISSN 2211-1247 . PMC 4555872 . PMID 26051934 .
- ^ де Taeye, Стивен В .; Озоровский, Габриэль; Торрентс-де-ла-Пенья, Альба; Гуттман, Миклош; Жюльен, Жан-Филипп; ван ден Керкхоф, Том LGM; Burger, Judith A .; Причард, Лаура К .; Пугач, Павел (17 декабря 2015 г.). «Иммуногенность стабилизированных тримеров оболочки ВИЧ-1 с пониженным воздействием ненейтрализующих эпитопов» . Cell . 163 (7): 1702–1715. DOI : 10.1016 / j.cell.2015.11.056 . ISSN 1097-4172 . PMC 4732737 . PMID 26687358 .
- ^ Уэлле DL, Плант I, Лэндри P, Барат C, Janelle ME, Flamand L, Трамбле MJ, благочинный P (апрель 2008). «Идентификация функциональных микроРНК, высвобождаемых посредством асимметричного процессинга TAR-элемента ВИЧ-1» . Исследования нуклеиновых кислот . 36 (7): 2353–65. DOI : 10.1093 / NAR / gkn076 . PMC 2367715 . PMID 18299284 .
- ^ KLASE Z, R Виноград, Дэвис Дж, Карпио л, Хилрает R, Heydarian М, фу S, Маккэффри Т, Meiri Е, Ayash-Рашковские М, Гилад S, Bentwich Z, Kashanchi F (2009). «МИРНК TAR ВИЧ-1 защищает от апоптоза, изменяя экспрессию клеточных генов» . Ретровирология . 6 (1): 18. DOI : 10,1186 / 1742-4690-6-18 . PMC 2654423 . PMID 19220914 .
- ^ Васудеван А.А., Смитс SH, Höppner А, Haussinger D, Коениг BW, Мунка С (ноябрь 2013 г. ). «Структурные особенности противовирусных ДНК цитидин дезаминаз» . Биологическая химия (Представленная рукопись). 394 (11): 1357–70. DOI : 10,1515 / HSZ-2013-0165 . PMID 23787464 . S2CID 4151961 .
- Перейти ↑ Garcia JV, Miller AD (апрель 1991 г.). "Серин-независимая от фосфорилирования подавление CD4 клеточной поверхности с помощью nef" . Природа . 350 (6318): 508–11. Bibcode : 1991Natur.350..508G . DOI : 10.1038 / 350508a0 . PMID 2014052 . S2CID 1628392 .
- Перейти ↑ Schwartz O, Maréchal V, Le Gall S, Lemonnier F, Heard JM (март 1996). «Эндоцитоз молекул класса I главного комплекса гистосовместимости индуцируется белком Nef ВИЧ-1» . Природная медицина . 2 (3): 338–42. DOI : 10.1038 / nm0396-338 . PMID 8612235 . S2CID 7461342 .
- ^ Stumptner-Cuvelette P, Morchoisne S, M Dugast, Le Gall S, Рапозо G, Schwartz O, Benaroch P (октябрь 2001). «Nef ВИЧ-1 нарушает презентацию антигена MHC класса II и поверхностную экспрессию» . Труды Национальной академии наук Соединенных Штатов Америки . 98 (21): 12144–9. Bibcode : 2001PNAS ... 9812144S . DOI : 10.1073 / pnas.221256498 . PMC 59782 . PMID 11593029 .
- ^ Аррильдт, Кэтрин Твигг; Джозеф, Сара Бет; Swanstrom, Рональд (март 2012). «Белок Env ВИЧ-1: разноцветный покров» . Текущие отчеты о ВИЧ / СПИДе . 9 (Текущие отчеты о ВИЧ / СПИДе): 53–63. DOI : 10.1007 / s11904-011-0107-3 . PMC 3658113 . PMID 22237899 .
- ^ a b Бергер Э.А., Домс Р.В., Фенё Э.М., Корбер Б.Т., Литтман Д.Р., Мур Дж. П., Саттентау К.Дж., Шуйтемейкер Х., Содроски Дж., Вайс Р.А. (1998). «Новая классификация ВИЧ-1» . Природа . 391 (6664): 240. Bibcode : 1998Natur.391..240B . DOI : 10.1038 / 34571 . PMID 9440686 . S2CID 2159146 .
- ^ a b c Coakley E, Petropoulos CJ, Whitcomb JM (2005). «Оценка использования корецептора ch vbgemokine при ВИЧ». Современное мнение об инфекционных заболеваниях . 18 (1): 9–15. DOI : 10.1097 / 00001432-200502000-00003 . PMID 15647694 . S2CID 30923492 .
- ^ Дэн Х, Лю Р., Эллмайер В., Чое С., Унутмаз Д., Буркхарт М., Ди Марцио П., Мармон С., Саттон Р. Э., Хилл К. М., Дэвис С. Б., Пайпер СК, Шалл Т. Дж., Литтман Д. Р., Ландау Н. Р. (1996). «Идентификация основного корецептора для первичных изолятов ВИЧ-1» . Природа . 381 (6584): 661–6. Bibcode : 1996Natur.381..661D . DOI : 10.1038 / 381661a0 . PMID 8649511 . S2CID 37973935 .
- Перейти ↑ Feng Y, Broder CC, Kennedy PE, Berger EA (1996). «Входной кофактор ВИЧ-1: функциональное клонирование кДНК семимембранного рецептора, сопряженного с G-белком» . Наука . 272 (5263): 872–7. Bibcode : 1996Sci ... 272..872F . DOI : 10.1126 / science.272.5263.872 . PMC 3412311 . PMID 8629022 . S2CID 44455027 .
- ^ Knight SC, Macatonia SE, Patterson S (1990). «ВИЧ-инфекция дендритных клеток». Международный обзор иммунологии . 6 (2–3): 163–75. DOI : 10.3109 / 08830189009056627 . PMID 2152500 .
- ^ Tang J, Kaslow RA (2003). «Влияние генетики хозяина на ВИЧ-инфекцию и прогрессирование заболевания в эпоху высокоактивной антиретровирусной терапии». СПИД . 17 (Дополнение 4): S51 – S60. DOI : 10.1097 / 00002030-200317004-00006 . PMID 15080180 .
- ↑ Zhu T, Mo H, Wang N, Nam DS, Cao Y, Koup RA, Ho DD (1993). «Генотипическая и фенотипическая характеристика больных ВИЧ-1 с первичной инфекцией». Наука . 261 (5125): 1179–81. Bibcode : 1993Sci ... 261.1179Z . DOI : 10.1126 / science.8356453 . PMID 8356453 .
- ^ van't Wout AB, Kootstra NA, Mulder-Kampinga GA, Albrecht-van Lent N, Scherpbier HJ, Veenstra J, Boer K, Coutinho RA, Miedema F, Schuitemaker H (1994). «Макрофаготропные варианты инициируют инфицирование вирусом иммунодефицита человека типа 1 после половой, парентеральной и вертикальной передачи» . Журнал клинических исследований . 94 (5): 2060–7. DOI : 10.1172 / JCI117560 . PMC 294642 . PMID 7962552 .
- ↑ Zhu T, Wang N, Carr A, Nam DS, Moor-Jankowski R, Cooper DA, Ho DD (1996). «Генетическая характеристика вируса иммунодефицита человека типа 1 в крови и генитальных выделениях: доказательства компартментализации и отбора вирусов во время передачи половым путем» . Журнал вирусологии . 70 (5): 3098–107. DOI : 10,1128 / JVI.70.5.3098-3107.1996 . PMC 190172 . PMID 8627789 .
- ^ Clevestig Р, Малькович я, Каспер С, Carlenor Е, Линдгрен s, NAVER л, Болин А.Б., Fenyő Е.М., Leitner Т, Ehrnst А (2005). «Фенотип X4 ВИЧ-типа 1 происходит от R5 у двух детей матерей, носящих X4, и не связан с передачей». Исследования СПИДа и ретровирусы человека . 21 (5): 371–8. DOI : 10,1089 / aid.2005.21.371 . PMID 15929699 .
- ^ Мур JP (1997). «Корецепторы: значение для патогенеза и терапии ВИЧ». Наука . 276 (5309): 51–2. DOI : 10.1126 / science.276.5309.51 . PMID 9122710 . S2CID 33262844 .
- ^ Карлссон А, Parsmyr К, Aperia К, Сандстрёе Е, Fenyő Е.М., Albert J (1994). «Тропизм клеток МТ-2 изолятов вируса иммунодефицита человека типа 1 как маркер ответа на лечение и развития лекарственной устойчивости». Журнал инфекционных болезней . 170 (6): 1367–75. DOI : 10.1093 / infdis / 170.6.1367 . PMID 7995974 .
- ^ Кут М, ван «т Wout А.Б., Kootstra Н.А., де GOEDE RE, Tersmette M, Schuitemaker H (1996). «Связь между изменениями клеточной нагрузки, эволюцией вирусного фенотипа и клональным составом вирусных популяций в ходе инфекции вируса иммунодефицита человека 1 типа» . Журнал инфекционных болезней . 173 (2): 349–54. DOI : 10.1093 / infdis / 173.2.349 . PMID 8568295 .
- Перейти ↑ Cheney K, McKnight A (2010). «Тропизм и болезнь ВИЧ-2». Лентивирусы и макрофаги: молекулярные и клеточные взаимодействия . Caister Academic Press . ISBN 978-1-904455-60-8.[ требуется страница ]
- ^ Б с д е е г Ch DC, Ким PS (1998). «Поступление ВИЧ и его подавление». Cell . 93 (5): 681–4. DOI : 10.1016 / S0092-8674 (00) 81430-0 . PMID 9630213 . S2CID 10544941 .
- ^ Б с д е е Wyatt R, Sodroski J (1998). «Гликопротеины оболочки ВИЧ-1: фузогены, антигены и иммуногены». Наука . 280 (5371): 1884–8. Bibcode : 1998Sci ... 280.1884W . DOI : 10.1126 / science.280.5371.1884 . PMID 9632381 .
- ^ а б Артос Дж., Цикала С., Мартинелли Э, Маклауд К., Ван Рик Д., Вей Д., Сяо З., Венстра Т. Д., Конрад Т. П., Лемпицки Р. А., Маклафлин С., Паскуччио М., Гопаул Р., Макналли Дж., Круз С. К., Цензоплано Н., Чунг Э., Рейтано К.Н., Коттилил С., Гуд Д.Д., Фаучи А.С. (2008). «Белок оболочки ВИЧ-1 связывается и передает сигнал через интегрин альфа (4) бета (7), рецептор самонаведения слизистой оболочки кишечника для периферических Т-клеток» . Иммунология природы . 9 (3): 301–9. DOI : 10.1038 / ni1566 . PMID 18264102 . S2CID 205361178 .
- ^ а б Папа М, Хаазе А.Т. (2003). «Передача, острая инфекция ВИЧ-1 и поиск стратегий предотвращения инфекции» . Природная медицина . 9 (7): 847–52. DOI : 10.1038 / nm0703-847 . PMID 12835704 . S2CID 26570505 .
- ^ Haedicke J, Brown C, Naghavi MH (август 2009). «Специфический для мозга фактор FEZ1 является определяющим фактором восприимчивости нейронов к инфекции ВИЧ-1» . Труды Национальной академии наук . 106 (33): 14040–14045. Bibcode : 2009PNAS..10614040H . DOI : 10.1073 / pnas.0900502106 . PMC 2729016 . PMID 19667186 .
- ^ Daecke Дж, Fackler ОТ, Dittmar МТ, Kräusslich HG (2005). «Участие клатрин-опосредованного эндоцитоза в проникновении вируса иммунодефицита человека типа 1» . Журнал вирусологии . 79 (3): 1581–1594. DOI : 10.1128 / jvi.79.3.1581-1594.2005 . PMC 544101 . PMID 15650184 .
- ^ Мияучи К, Ким У, Latinovic О, Морозов В, Меликян ГБ (2009 г.). «ВИЧ проникает в клетки посредством эндоцитоза и динамин-зависимого слияния с эндосомами» . Cell . 137 (3): 433–444. DOI : 10.1016 / j.cell.2009.02.046 . PMC 2696170 . PMID 19410541 .
- ^ Кох Р, Лампа М, Годинес WJ, Мюллер В, Рор К, Kräusslich HG, Леман МДж (2009). «Визуализация слияния псевдотипированных частиц ВИЧ-1 в реальном времени с помощью микроскопии живых клеток» . Ретровирология . 6 : 84. DOI : 10,1186 / 1742-4690-6-84 . PMC 2762461 . PMID 19765276 .
- ^ Торли JA, McKeating JA, Раппопорт JZ (2010). «Механизм проникновения вируса: крадется в парадную дверь» . Протоплазма . 244 (1–4): 15–24. DOI : 10.1007 / s00709-010-0152-6 . PMC 3038234 . PMID 20446005 .
- ^ Перманьер M, Баллан E, Эст JA (2010). «Эндоцитоз ВИЧ: все идет». Тенденции в микробиологии . 18 (12): 543–551. DOI : 10.1016 / j.tim.2010.09.003 . PMID 20965729 .
- ^ a b c Чжэн Ю.Х., Ловсин Н., Петерлин Б.М. (2005). «Недавно идентифицированные факторы хозяина модулируют репликацию ВИЧ». Письма иммунологии . 97 (2): 225–34. DOI : 10.1016 / j.imlet.2004.11.026 . PMID 15752562 .
- ^ «IV. Вирусы> F. Жизненные циклы вирусов животных> 3. Жизненный цикл ВИЧ» . Домашняя страница микробиологии Дока Кайзера . Общественный колледж округа Балтимор. Январь 2008. Архивировано из оригинала 26 июля 2010 года.
- ^ Hiscott Дж, Kwon Н, Р Genin (2001). «Враждебные поглощения: вирусное присвоение пути NF-kB» . Журнал клинических исследований . 107 (2): 143–151. DOI : 10.1172 / JCI11918 . PMC 199181 . PMID 11160127 .
- ^ Поллард VW, Малим MH (1998). «Белок HIV-1 Rev». Ежегодный обзор микробиологии . 52 : 491–532. DOI : 10.1146 / annurev.micro.52.1.491 . PMID 9891806 .
- ^ Butsch, M .; Борис-Лори, К. (2002). «Судьба несплицированной ретровирусной РНК: рибосома и / или вирион?» . Журнал вирусологии . 76 (7): 3089–94. DOI : 10,1128 / JVI.76.7.3089-3094.2002 . PMC 136024 . PMID 11884533 .
- ^ Хельмунд, Крис; Рычаг, Эндрю М.Л. (14 июля 2016 г.). «Координация упаковки геномной РНК с вирусной сборкой в ВИЧ-1» . Вирусы . 8 (7): 192. DOI : 10,3390 / v8070192 . ISSN 1999-4915 . PMC 4974527 . PMID 27428992 .
- ^ Сото-Рифо, Р .; Лимузен, Т .; Рубилар, PS; Ricci, EP; Décimo, D .; Moncorgé, O .; Trabaud, MA; André, P .; Cimarelli, A .; Ольманн, Т. (2011). «Различные эффекты структуры TAR на трансляцию геномных РНК ВИЧ-1 и ВИЧ-2» . Исследования нуклеиновых кислот . 40 (6): 2653–2667. DOI : 10.1093 / NAR / gkr1093 . PMC 3315320 . PMID 22121214 .
- ^ Саад, Джамиль С .; Мурио, Дельфин М. (28 июля 2015 г.). Роль липидов в сборке вирусов . Frontiers Media SA. ISBN 9782889195824.
- ^ Риччи, EP; Эрбрето, Швейцария; Decimo, D .; Schaupp, A .; Датта, SAK; Rein, A .; Darlix, J. -L .; Ольманн, Т. (2008). «Экспрессия геномной РНК ВИЧ-2 in vitro контролируется тремя отдельными внутренними входными сегментами рибосомы, которые регулируются протеазой ВИЧ и полипротеином Gag» . РНК . 14 (7): 1443–55. DOI : 10,1261 / rna.813608 . PMC 2441975 . PMID 18495939 .
- ^ а б Ху WS, Темин Х.М. (1990). «Ретровирусная рекомбинация и обратная транскрипция». Наука . 250 (4985): 1227–33. Bibcode : 1990Sci ... 250.1227H . DOI : 10.1126 / science.1700865 . PMID 1700865 .
- ^ a b Шарпантье C, Нора T, Tenaillon O, Клавель F, Hance AJ (2006). «Обширная рекомбинация квазивидов вируса иммунодефицита человека типа 1 вносит важный вклад в вирусное разнообразие у отдельных пациентов» . Журнал вирусологии . 80 (5): 2472–82. DOI : 10,1128 / JVI.80.5.2472-2482.2006 . PMC 1395372 . PMID 16474154 .
- ^ Нора Т, Шарпантье С, Tenaillon О, Hoede С, Клавел Ж, Hance AJ (2007). «Вклад рекомбинации в эволюцию вирусов иммунодефицита человека, выражающих устойчивость к антиретровирусному лечению» . Журнал вирусологии . 81 (14): 7620–8. DOI : 10,1128 / JVI.00083-07 . PMC 1933369 . PMID 17494080 .
- Перейти ↑ Chen J, Powell D, Hu WS (2006). «Высокая частота генетической рекомбинации - общая черта репликации лентивирусов приматов» . Журнал вирусологии . 80 (19): 9651–8. DOI : 10,1128 / JVI.00936-06 . PMC 1617242 . PMID 16973569 .
- ^ a b Bonhoeffer S, Chappey C, Parkin NT, Whitcomb JM, Petropoulos CJ (2004). «Доказательства положительного эпистаза при ВИЧ-1». Наука . 306 (5701): 1547–50. Bibcode : 2004Sci ... 306.1547B . DOI : 10.1126 / science.1101786 . PMID 15567861 . S2CID 45784964 .
- ^ Israël N, Гужеро-Pocidalo М.А. (1997). «Окислительный стресс при инфекции вируса иммунодефицита человека» . Клеточные и молекулярные науки о жизни . 53 (11–12): 864–70. DOI : 10.1007 / s000180050106 . PMID 9447238 . S2CID 22663454 .
- ^ Michod RE Бернштейн H, Nedelcu AM (май 2008). «Адаптивное значение пола у микробных патогенов» (PDF) . Инфекция, генетика и эволюция . 8 (3): 267–85. DOI : 10.1016 / j.meegid.2008.01.002 . PMID 18295550 .
- ^ Hallenberger S, Бош В, Angliker Н, Шоу Е, Klenk HD, Гартен Вт (26 ноября, 1992 г.). «Ингибирование фурин-опосредованной активации расщепления гликопротеина gp160 ВИЧ-1» . Природа . 360 (6402): 358–61. Bibcode : 1992Natur.360..358H . DOI : 10.1038 / 360358a0 . PMID 1360148 . S2CID 4306605 .
- ^ Gelderblom HR (1997). «Тонкая структура ВИЧ и SIV» (PDF) . В Лос-Аламосской национальной лаборатории (ред.). Сборник последовательностей ВИЧ . Лос-Аламосская национальная лаборатория . С. 31–44.
- ^ а б в г Чжан С., Чжоу С., Гроппелли Э, Пеллегрино П., Уильямс И., Броу П., Цепь Б. М., Веселый С. (2015). «Гибридные механизмы распространения и активация Т-лимфоцитов определяют динамику инфицирования ВИЧ-1» . PLOS Вычислительная биология . 11 (4): e1004179. arXiv : 1503.08992 . Bibcode : 2015PLSCB..11E4179Z . DOI : 10.1371 / journal.pcbi.1004179 . PMC 4383537 . PMID 25837979 .
- ^ a b Веселый C, Kashefi K, Hollinshead M, Sattentau QJ (2004). «Перенос клеток ВИЧ-1 в клетки через Env-индуцированный актин-зависимый синапс» . Журнал экспериментальной медицины . 199 (2): 283–293. DOI : 10,1084 / jem.20030648 . PMC 2211771 . PMID 14734528 .
- ^ Sattentau Q (2008). «Избегая пустоты: распространение вирусов от клетки к клетке» . Обзоры природы микробиологии . 6 (11): 815–826. DOI : 10.1038 / nrmicro1972 . PMID 18923409 . S2CID 20991705 .
- ^ Дункан CJ, Рассел Р., Sattentau QJ (2013). «Высокая множественность передачи ВИЧ-1 от клетки к клетке от макрофагов к CD4 + Т-клеткам ограничивает антиретровирусную эффективность» . СПИД . 27 (14): 2201–2206. DOI : 10.1097 / QAD.0b013e3283632ec4 . PMC 4714465 . PMID 24005480 .
- ^ Sewald X, Гонсалес DG, Haberman AM, Mothes W (2012). «Визуализация вирусологических синапсов in vivo» . Nature Communications . 3 : 1320. Bibcode : 2012NatCo ... 3.1320S . DOI : 10.1038 / ncomms2338 . PMC 3784984 . PMID 23271654 .
- Перейти ↑ Sigal A, Kim JT, Balazs AB, Dekel E, Mayo A, Milo R, Baltimore D (2011). «Распространение ВИЧ от клетки к клетке позволяет продолжать репликацию, несмотря на антиретровирусную терапию» . Природа . 477 (7362): 95–98. Bibcode : 2011Natur.477 ... 95S . DOI : 10,1038 / природа10347 . PMID 21849975 . S2CID 4409389 .
- ^ a b c d Робертсон Д. Л., Хан Б. Х., Sharp PM (1995). «Рекомбинация в вирусах СПИДа» . Журнал молекулярной эволюции . 40 (3): 249–59. Bibcode : 1995JMolE..40..249R . DOI : 10.1007 / BF00163230 . PMID 7723052 . S2CID 19728830 .
- ^ Rambaut A, D Посада, Crandall KA, Холмс EC (январь 2004). «Причины и последствия эволюции ВИЧ» . Природа Обзоры Генетики . 5 (52–61): 52–61. DOI : 10.1038 / nrg1246 . PMID 14708016 . S2CID 5790569 .
- ^ Perelson AS, Ribeiro RM (октябрь 2008). «Оценка эффективности лекарств и вирусных динамических параметров: ВИЧ и ВГС» . Статистика в медицине . 27 (23): 4647–57. DOI : 10.1002 / sim.3116 . PMID 17960579 .
- ^ a b Sodora DL, Allan JS, Apetrei C, Brenchley JM, Douek DC, Else JG, Estes JD, Hahn BH, Hirsch VM, Kaur A, Kirchhoff F, Muller-Trutwin M, Pandrea I, Schmitz JE, Silvestri G ( 2009 г.). «К вакцине против СПИДа: уроки естественного заражения вирусом обезьяньего иммунодефицита африканских приматов-хозяев» . Природная медицина . 15 (8): 861–865. DOI : 10.1038 / nm.2013 . PMC 2782707 . PMID 19661993 .
- ^ Holzammer S, Holznagel Е, Каул А, Курта R, S Norley (2001). «Высокие вирусные нагрузки у африканских зеленых мартышек, инфицированных естественным и экспериментальным путем SIVagm». Вирусология . 283 (2): 324–31. DOI : 10.1006 / viro.2001.0870 . PMID 11336557 .
- ^ Kurth, R .; Норли, С. (1996). «Почему у естественных носителей ВИО не развивается обезьяний СПИД?». Журнал исследований NIH . 8 : 33–37.
- ^ Байера М, Dittmar МТ, Cichutek К, Р Курта (1991). «Развитие vivo генетической изменчивости вируса иммунодефицита обезьян» . Труды Национальной академии наук Соединенных Штатов Америки . 88 (18): 8126–30. Bibcode : 1991PNAS ... 88.8126B . DOI : 10,1073 / пнас.88.18.8126 . PMC 52459 . PMID 1896460 .
- ^ Дэниел Мэриленд, Король NW, Летвин Н.Л., Хант Р.Д., Сегал П.К., Дерозье RC (1984). «Новый ретровирус типа D, выделенный от макак с синдромом иммунодефицита». Наука . 223 (4636): 602–5. Bibcode : 1984Sci ... 223..602D . DOI : 10.1126 / science.6695172 . PMID 6695172 .
- ^ a b Кил Б. Ф., Джонс Дж. Х., Терио К. А., Эстес Дж. Д., Рудичелл Р. С., Уилсон М. Л., Ли Y, ЖЖ Г. Х., Бизли Т. М., Шумахер-Станки Дж., Вроблевски Е., Моссер А., Рафаэль Дж., Каменя С., Лонсдорф Е. В., Трэвис Д.А., Мленгея Т., Кинсел М.Дж., Остальное Дж. Г., Сильвестри Дж., Гудолл Дж., Шарп П.М., Шоу Г.М., Пьюзи А.Е., Хан Б.Х. (2009). «Повышенная смертность и СПИД-подобная иммунопатология у диких шимпанзе, инфицированных SIVcpz» . Природа . 460 (7254): 515–519. Bibcode : 2009Natur.460..515K . DOI : 10,1038 / природа08200 . PMC 2872475 . PMID 19626114 .
- ^ Schindler M, Münch J, Kutsch O, Li H, Santiago ML, Bibollet-Ruche F, Müller-Trutwin MC, Novembre FJ, Peeters M, Courgnaud V, Bailes E, Roques P, Sodora DL, Silvestri G, Sharp PM, Хан Б. Х., Кирхгоф Ф (2006). «Опосредованное Nef подавление активации Т-клеток было потеряно в лентивирусной линии, которая дала начало ВИЧ-1». Cell . 125 (6): 1055–67. DOI : 10.1016 / j.cell.2006.04.033 . PMID 16777597 . S2CID 15132918 .
- Перейти ↑ Thomson MM, Pérez-Alvarez L, Nájera R (2002). «Молекулярная эпидемиология генетических форм ВИЧ-1 и ее значение для разработки вакцин и терапии». Ланцетные инфекционные болезни . 2 (8): 461–471. DOI : 10.1016 / S1473-3099 (02) 00343-2 . PMID 12150845 .
- ^ Карр Ю.К., Фоль ЕТ, Leitner Т, Салминны М, Корбер В, McCutchan F (1998). «Контрольные последовательности, представляющие основное генетическое разнообразие ВИЧ-1 в условиях пандемии» (PDF) . В Лос-Аламосской национальной лаборатории (ред.). Сборник последовательностей ВИЧ . Лос-Аламос, Нью-Мексико : Национальная лаборатория Лос-Аламоса . С. 10–19.
- ^ Османов S, Pattou С, Н Уокер, Schwardländer В, Эспарза J (2002). «Предполагаемое глобальное распространение и региональное распространение генетических подтипов ВИЧ-1 в 2000 году». Журнал синдромов приобретенного иммунодефицита . 29 (2): 184–190. DOI : 10.1097 / 00042560-200202010-00013 . PMID 11832690 . S2CID 12536801 .
- ^ Перрин L, L Kaiser, Yerly S (2003). «Путешествие и распространение генетических вариантов ВИЧ-1». Ланцетные инфекционные болезни . 3 (1): 22–27. DOI : 10.1016 / S1473-3099 (03) 00484-5 . PMID 12505029 .
- ^ a b Plantier JC, Леоз М, Дикерсон Дж., Де Оливейра Ф, Кордонье Ф, Леме В., Дамонд Ф, Робертсон Д. Л., Саймон Ф (август 2009 г.). «Новый вирус иммунодефицита человека, полученный от горилл» . Природная медицина . 15 (8): 871–2. DOI : 10.1038 / nm.2016 . PMID 19648927 . S2CID 76837833 . Выложите резюме .
- ^ Кил Б. Ф., Ван Heuverswyn F, Li Y, Bailes Е, Такехис J, Сантьяго М.Л., Bibollet-Рюш Ж, Чен Y, Вайн Л.В., Liegeois F, Loul S, Ngole Е.М., Bienvenue Y, Делапорт Е, Брукфильд ДФ, Sharp PM, Шоу Г.М., Питерс М., Хан Б.Х. (28 июля 2006 г.). «Шимпанзе - резервуары пандемического и непандемического ВИЧ-1» . Наука . 313 (5786): 523–6. Bibcode : 2006Sci ... 313..523K . DOI : 10.1126 / science.1126531 . PMC 2442710 . PMID 16728595 .
- ^ a b c d Kumaranayake, L .; Уоттс, К. (2001). «Распределение ресурсов и установление приоритетов мероприятий по ВИЧ / СПИДу: борьба с генерализованной эпидемией в странах Африки к югу от Сахары». Журнал международного развития . 13 (4): 451–466. DOI : 10.1002 / jid.797 .
- ^ Кляйнман S (сентябрь 2004). «Информация для пациентов: сдача и переливание крови» . Своевременно. Архивировано из оригинального 12 апреля 2008 года.
- ^ a b Центры по контролю и профилактике заболеваний (2001). «Пересмотренное руководство по консультированию, тестированию и направлению на ВИЧ». Рекомендации и отчеты MMWR . 50 (RR – 19): 1–57. PMID 11718472 .
- ^ Celum CL, Кумбс RW, Лэфферти Вт, Инуи TS, Луи PH, Ворота СА, McCreedy BJ, Иган Р, Т - Гроув, Александр С (1991). «Неопределенный вестерн-блоттинг вируса иммунодефицита человека 1 типа: риск сероконверсии, специфичность дополнительных тестов и алгоритм оценки». Журнал инфекционных болезней . 164 (4): 656–664. DOI : 10.1093 / infdis / 164.4.656 . PMID 1894929 .
- ^ "Сравнение стран :: ВИЧ / СПИД - Смерти" . The World Factbook, Центральное разведывательное управление.
- ^ Чоу, Роджер; Селф, Шелли; Дана, Трейси; Бугацос, Кристина; Захер, Бернадетт; Блазина, Ян; Кортуис, П. Тодд (20 ноября 2012 г.). «Скрининг на ВИЧ: систематический обзор для обновления рекомендаций Целевой группы по профилактическим услугам США 2005 г.» . Анналы внутренней медицины . 157 (10): 706–718. DOI : 10.7326 / 0003-4819-157-10-201211200-00007 . ISSN 1539-3704 . PMID 23165662 . S2CID 27494096 .
- ^ Chou R, Хаффман LH, Fu R, Смитс AK, Korthuis PT (июль 2005). «Скрининг на ВИЧ: обзор доказательств для Целевой группы США по профилактическим услугам» . Анналы внутренней медицины . 143 (1): 55–73. DOI : 10.7326 / 0003-4819-143-1-200507050-00010 . PMID 15998755 .
- Перейти ↑ Tolle MA, Schwarzwald HL (15 июля 2010 г.). «Постконтактная профилактика вируса иммунодефицита человека». Американский семейный врач . 82 (2): 161–6. PMID 20642270 .
- ^ «Краткое справочное руководство - Лабораторное тестирование для диагностики ВИЧ-инфекции: обновленные рекомендации» (PDF) . cdc.gov . Департамент здравоохранения штата Нью-Йорк. 27 июня 2014 г. С. 1–2. Архивировано из оригинального (PDF) 2 марта 2017 года . Проверено 13 апреля 2017 года .
- ^ «Лечение ВИЧ: одобренные FDA лекарства против ВИЧ» . AIDSinfo.
- ^ Швейцарская национальная комиссия по СПИДу (15 октября 2016 г.). «Швейцарское заявление» . ВИЧ i-Base . Проверено 2 апреля 2019 года .
- ^ Вернацца, P; Бернар, Э.Дж. (29 января 2016 г.). «ВИЧ не передается при полностью подавляющей терапии: Швейцарское заявление - восемь лет спустя» . Швейцарский медицинский еженедельник . 146 : w14246. DOI : 10.4414 / smw.2016.14246 . PMID 26824882 .
- ^ The Lancet HIV (ноябрь 2017 г.). «U = U взлет в 2017 году» . От редакции. Ланцет ВИЧ . 44 (11): e475. DOI : 10.1016 / S2352-3018 (17) 30183-2 . PMID 29096785 .
- ^ "Не могу передать это дальше" . Терренс Хиггинс Trust . 2019. Архивировано 7 апреля 2019 года . Проверено 2 апреля 2019 года .
- ^ Бавингтон, Бенджамин Р .; Пинто, Энджи Н .; Пханупхак, Ниттая; Гринштейн, Беатрис; Prestage, Garrett P .; Заблоцкая-Манос Ирина Б .; и другие. (2018). «Подавление вируса и передача ВИЧ в серодискордантных мужских парах: международное проспективное обсервационное когортное исследование». Ланцет ВИЧ . 5 (8): e438 – e447. DOI : 10.1016 / S2352-3018 (18) 30132-2 . PMID 30025681 .
- ^ Роджер, Элисон Дж .; Камбиано, Валентина; Бруун, Тина; Вернацца, Пьетро; Коллинз, Саймон; ван Лунзен, Ян; и другие. (2016). «Сексуальная активность без презервативов и риск передачи ВИЧ в серодифферентных парах, когда ВИЧ-положительный партнер использует подавляющую антиретровирусную терапию» . JAMA . 316 (2): 171–181. DOI : 10,1001 / jama.2016.5148 . PMID 27404185 .
- ^ Rodger, A. (для PARTNER исследовательской группы) (июль 2018). Риск передачи ВИЧ при сексе без презерватива в парах МСМ с подавляющей АРТ: исследование PARTNER2 расширило результаты среди геев . AIDS2018: 22-я Международная конференция по СПИДу. Амстердам, Нидерланды . Проверено 2 апреля 2019 года .
- ↑ a b c Хоффман, Хиллари (10 января 2019 г.). «Наука ясна: с ВИЧ неопределяемое равно непередаваемое» (пресс-релиз). Национальные институты здоровья . Национальный институт аллергии и инфекционных заболеваний . Проверено 3 мая 2019 года .
Директор NIAID Энтони С. Фаучи, доктор медицины, и его коллеги обобщают результаты крупных клинических испытаний и когортных исследований, подтверждающих, что U = U. Знаковое клиническое исследование HPTN 052, финансируемое Национальным институтом здравоохранения, показало, что между серодифферентными гетеросексуальными парами ВИЧ не было связанной передачи ВИЧ, когда у партнера, живущего с ВИЧ, вирусная нагрузка была длительно подавлена. Впоследствии исследования PARTNER и Opposites Attract подтвердили эти выводы и распространили их на пары мужчина-мужчина. ... Успех U = U как метода профилактики ВИЧ зависит от достижения и поддержания неопределяемой вирусной нагрузки путем ежедневного приема АРТ в соответствии с предписаниями.
- ^ Коэн, Майрон С .; Chen, Ying Q .; Макколи, Мэрибет; Гэмбл, Тереза; Hosseinipour, Mina C .; Кумарасами, Нагалингесваран; и другие. (2016). «Антиретровирусная терапия для профилактики передачи ВИЧ-1» . N. Engl. J. Med. 375 (9): 830–839. DOI : 10.1056 / NEJMoa1600693 . PMC 5049503 . PMID 27424812 .
- ↑ Ходсон, Мэтью (17 ноября 2017 г.). U = U: Разговор с пациентами о риске передачи (PDF) . Осенняя конференция Британской ассоциации по ВИЧ 2017 . Проверено 3 мая 2019 года . ( аннотация для презентации от имени NAM / aidsmap )
- ^ «Консенсусное заявление: риск передачи ВИЧ половым путем от человека, живущего с ВИЧ, у которого вирусная нагрузка не поддается обнаружению» . Кампания по предотвращению доступа . 21 июля 2016 . Проверено 2 апреля 2019 года . Примечание . На момент получения заявления и списка одобрений он последний раз обновлялся 23 августа 2018 г. и включал «более 850 организаций из почти 100 стран».
- ^ Boseley, Сара; Девлин, Ханна (3 мая 2019 г.). «Конец СПИДу на горизонте, поскольку огромное исследование показало, что наркотики останавливают передачу ВИЧ» . Хранитель . Проверено 3 мая 2019 года .
- ^ Чжу, Цзя; Хладик, Флориан; Вудворд, Аманда; Клок, Алексис; Пэн, Дао; Джонстон, Кристина; Ремингтон, Майкл; Магарет, Амалия; Koelle, Дэвид М .; Уолд, Анна; Кори, Лоуренс (август 2009 г.). «Сохранение рецептор-положительных клеток ВИЧ-1 после реактивации ВПГ-2: потенциальный механизм увеличения приобретения ВИЧ-1» . Природная медицина . 15 (8): 886–892. DOI : 10.1038 / nm.2006 . ISSN 1078-8956 . PMC 2723183 . PMID 19648930 .
- ^ Looker, Кэтрин Дж .; Elmes, Jocelyn AR; Gottlieb, Sami L .; Шиффер, Джошуа Т .; Викерман, Питер; Тернер, Кэтрин ME; Бойли, Мари-Клод (декабрь 2017 г.). «Влияние инфекции HSV-2 на последующее заражение ВИЧ: обновленный систематический обзор и метаанализ» . Ланцет. Инфекционные болезни . 17 (12): 1303–1316. DOI : 10.1016 / S1473-3099 (17) 30405-X . ISSN 1474-4457 . PMC 5700807 . PMID 28843576 .
- ^ «В этот день». Новости и записи . 18 мая 2020 г. с. 2А.
- ^ Mandell, Джеральд L .; Беннетт, Джон Э .; Долин, Рафаэль, ред. (2010). «Глава 169». Принципы и практика инфекционных болезней Манделла, Дугласа и Беннета (7-е изд.). Филадельфия: Черчилль Ливингстон / Эльзевьер. ISBN 978-0-443-06839-3.[ требуется страница ]
- Перейти ↑ Gottlieb MS (2006). «Пневмоцистная пневмония - Лос-Анджелес. 1981» . Американский журнал общественного здравоохранения . 96 (6): 980–1, обсуждение 982–3. DOI : 10,2105 / AJPH.96.6.980 . PMC 1470612 . PMID 16714472 . Архивировано 22 апреля 2009 года.
- ↑ Friedman-Kien AE (октябрь 1981 г.). «Синдром распространенной саркомы Капоши у молодых гомосексуалистов». Журнал Американской академии дерматологии . 5 (4): 468–71. DOI : 10.1016 / S0190-9622 (81) 80010-2 . PMID 7287964 .
- ^ Хаймс KB, Cheung T, Greene JB, Проза Н.С., Marcus A, Ballard H, Уильям DC, Laubenstein LJ (сентябрь 1981). «Саркома Капоши у гомосексуалистов - сообщение о восьми случаях» . Ланцет . 2 (8247): 598–600. DOI : 10.1016 / S0140-6736 (81) 92740-9 . PMID 6116083 . S2CID 43529542 .
- ^ a b Basavapathruni A, Андерсон К.С. (декабрь 2007 г.). «Обратная транскрипция пандемии ВИЧ-1». Журнал FASEB . 21 (14): 3795–3808. DOI : 10,1096 / fj.07-8697rev . PMID 17639073 . S2CID 24960391 .
- ^ Ледерберг, Джошуа, изд. (2000). Энциклопедия микробиологии (2-е изд.). Берлингтон: Эльзевир. п. 106. ISBN 978-0-08-054848-7. Проверено 9 июня, 2016 .
- ^ Центры по контролю заболеваний (1982). «Стойкая генерализованная лимфаденопатия среди мужчин-гомосексуалистов» . Еженедельный отчет о заболеваемости и смертности . 31 (19): 249–251. PMID 6808340 .
- ^ Барре-Синусси Ж, Chermann JC, Rey F, Nugeyre МТ, Chamaret S, Gruest Дж, Dauguet С, Axler-Blin С, Vézinet-Брун Ж, Rouzioux С, Розенбаум Вт, Монтанье L (1983). «Выделение Т-лимфотропного ретровируса от пациента с риском синдрома приобретенного иммунодефицита (СПИД)» . Наука . 220 (4599): 868–871. Bibcode : 1983Sci ... 220..868B . DOI : 10.1126 / science.6189183 . PMID 6189183 . S2CID 390173 .
- ^ a b Центры по контролю за заболеваниями (1982). «Оппортунистические инфекции и саркома Капоши среди гаитян в США» . Еженедельный отчет о заболеваемости и смертности . 31 (26): 353–354, 360–361. PMID 6811853 .
- ↑ Altman LK (11 мая 1982 г.). «Новое гомосексуальное расстройство беспокоит чиновников здравоохранения» . Нью-Йорк Таймс . Проверено 31 августа 2011 года .
- Перейти ↑ Gilman, Sander L (1987). Гилман, Сандер Л. (ред.). «СПИД и сифилис: иконография болезней». Октябрь . 43 : 87–107. DOI : 10.2307 / 3397566 . JSTOR 3397566 .
- ^ "Продвигаясь вперед в адских обстоятельствах" (PDF) . Американская ассоциация развития науки . 28 июля 2006 . Проверено 23 июня 2008 года .
- ↑ Kher U (27 июля 1982 г.). «Имя чумы» . Время . Архивировано 7 марта 2008 года . Проверено 10 марта 2008 года .
- ^ Центры по контролю заболеваний (1982). «Последняя информация о синдроме приобретенного иммунодефицита (СПИД) - США». Еженедельный отчет о заболеваемости и смертности . 31 (37): 507–508, 513–514. PMID 6815471 .
- ↑ Gallo RC, Sarin PS, Gelmann EP, Robert-Guroff M, Richardson E, Kalyanaraman VS, Mann D, Sidhu GD, Stahl RE, Zolla-Pazner S, Leibowitch J, Popovic M (1983). «Выделение вируса Т-клеточного лейкоза человека при синдроме приобретенного иммунодефицита (СПИД)». Наука . 220 (4599): 865–867. Bibcode : 1983Sci ... 220..865G . DOI : 10.1126 / science.6601823 . PMID 6601823 .
- ^ Барре-Синусси Ж, Chermann JC, Rey F, Nugeyre МТ, Chamaret S, Gruest Дж, Dauguet С, Axler-Blin С, Vézinet-Брун Ж, Rouzioux С, Розенбаум Вт, Монтанье L (1983). «Выделение Т-лимфотропного ретровируса от пациента с риском синдрома приобретенного иммунодефицита (СПИД)» . Наука . 220 (4599): 868–871. Bibcode : 1983Sci ... 220..868B . DOI : 10.1126 / science.6189183 . PMID 6189183 . S2CID 390173 .
- ^ «Нобелевская премия по физиологии и медицине 2008 года - пресс-релиз» . www.nobelprize.org . Проверено 28 января 2018 года .
- ^ Crewdson, Джон (30 мая 1991). "ГАЛЛО ДОПУСКАЕТ ОБНАРУЖЕНИЕ ФРАНЦУЗСКОГО ВИРУСА СПИДА" . Чикаго Трибьюн . Проверено 25 апреля 2020 года .
- ^ Олдрич, Роберт; Уотерспун, Гарри, ред. (2001). Кто есть кто в истории геев и лесбиянок . Лондон: Рутледж. п. 154. ISBN 978-0-415-22974-6.
- ^ Леви JA; и другие. (1984). «Выделение лимфоцитопатических ретровирусов от больных СПИДом Сан-Франциско». Наука . 225 (4664): 840–842. Bibcode : 1984Sci ... 225..840L . DOI : 10.1126 / science.6206563 . PMID 6206563 .
- ↑ Леви Дж. А., Камински Л. С., Морроу В. Дж., Стеймер К., Люцив П., Дина Д., Хокси Дж., Оширо Л. (1985). «Заражение ретровирусом, связанное с синдромом приобретенного иммунодефицита». Анналы внутренней медицины . 103 (5): 694–699. DOI : 10.7326 / 0003-4819-103-5-694 . PMID 2996401 .
- Перейти ↑ Sharp PM, Hahn BH (2011). «Истоки ВИЧ и пандемия СПИДа» . Перспективы Колд-Спринг-Харбор в медицине . 1 (1): a006841. DOI : 10.1101 / cshperspect.a006841 . PMC 3234451 . PMID 22229120 .
- ^ Фариа Н.Р., Rambaut А, Сушар М.А., Baele G, Бедфорд Т, Уорд МДж, Татэм AJ, Соуза JD, Arinaminpathy N, Pépin J, D Посада, Петерс М, Pybus О.Г., Lemey Р (2014). «Раннее распространение и возгорание эпидемии ВИЧ-1 в человеческих популяциях» . Наука . 346 (6205): 56–61. Bibcode : 2014Sci ... 346 ... 56F . DOI : 10.1126 / science.1256739 . PMC 4254776 . PMID 25278604 .
- ^ Гао Ф, Бейлс Э, Робертсон Д.Л., Чен Ю., Роденбург С.М., Майкл С.Ф., Камминс Л. «Происхождение ВИЧ-1 у шимпанзе Pan troglodytes troglodytes» . Природа . 397 (6718): 436–41. Bibcode : 1999Natur.397..436G . DOI : 10,1038 / 17130 . PMID 9989410 . S2CID 4432185 .
- ^ Кил Б. Ф., Ван Heuverswyn F, Li Y, Bailes Е, Такехис J, Сантьяго М.Л., Bibollet-Рюш Ж, Чен Y, Вайн Л.В., Liegeois F, Loul S, Ngole Е.М., Bienvenue Y, Делапорт Е, Брукфильд ДФ, Sharp PM, Шоу GM, Петерс М., Хан Б.Х. (2006). «Шимпанзе - резервуары пандемического и непандемического ВИЧ-1» . Наука . 313 (5786): 523–6. Bibcode : 2006Sci ... 313..523K . DOI : 10.1126 / science.1126531 . PMC 2442710 . PMID 16728595 .
- ^ Goodier JL, Kazazian HH (2008). «Ретротранспозоны снова: сдерживание и реабилитация паразитов». Cell . 135 (1): 23–35. DOI : 10.1016 / j.cell.2008.09.022 . PMID 18854152 . S2CID 3093360 .
- ↑ Sharp PM, Bailes E, Chaudhuri RR, Rodenburg CM, Santiago MO, Hahn BH (2001). «Истоки вирусов синдрома приобретенного иммунодефицита: где и когда?» . Философские труды Королевского общества B . 356 (1410): 867–76. DOI : 10.1098 / rstb.2001.0863 . PMC 1088480 . PMID 11405934 .
- ^ Калиш М. Л., Вольф Н. Д., Ndongmo СВ, McNicholl Дж, Роббинс К.Е., Аидоо М, Fonjungo ПН, Alemnji G, Zeh С, Джоко CF, Mpoudi-Ngole Е, Берк DS, Люди ТМ (2005). «Охотники из Центральной Африки, зараженные вирусом обезьяньего иммунодефицита» . Возникающие инфекционные заболевания . 11 (12): 1928–30. DOI : 10.3201 / eid1112.050394 . PMC 3367631 . PMID 16485481 .
- ^ a b Маркс PA, Alcabes PG, Drucker E (2001). «Серийный перенос вируса обезьяньего иммунодефицита через нестерильные инъекции и появление эпидемического вируса иммунодефицита человека в Африке» . Философские труды Королевского общества B . 356 (1410): 911–20. DOI : 10.1098 / rstb.2001.0867 . PMC 1088484 . PMID 11405938 .
- ^ Worobey M, Gemmel M, Teuwen DE, Haselkorn T, Kunstman K, Bunce M, Muyembe JJ, Kabongo JM, Kalengayi RM, Van Marck E, Gilbert MT, Wolinsky SM (2008). "Direct evidence of extensive diversity of HIV-1 in Kinshasa by 1960". Nature. 455 (7213): 661–4. Bibcode:2008Natur.455..661W. doi:10.1038/nature07390. PMC 3682493. PMID 18833279.
- ^ a b de Sousa JD, Müller V, Lemey P, Vandamme AM (2010). Martin DP (ed.). "High GUD incidence in the early 20th century created a particularly permissive time window for the origin and initial spread of epidemic HIV strains". PLOS ONE. 5 (4): e9936. Bibcode:2010PLoSO...5.9936S. doi:10.1371/journal.pone.0009936. PMC 2848574. PMID 20376191.
- ^ Zhu T, Korber BT, Nahmias AJ, Hooper E, Sharp PM, Ho DD (1998). "An African HIV-1 Sequence from 1959 and Implications for the Origin of the epidemic". Nature. 391 (6667): 594–7. Bibcode:1998Natur.391..594Z. doi:10.1038/35400. PMID 9468138. S2CID 4416837.
- ^ Kolata, Gina (October 28, 1987). "Boy's 1969 death suggests AIDS invaded U.S. several times". The New York Times. Retrieved February 11, 2009.
- ^ Chitnis A, Rawls D, Moore J (2000). "Origin of HIV type 1 in colonial French equatorial Africa?". AIDS Research and Human Retroviruses. 16 (1): 5–8. doi:10.1089/088922200309548. PMID 10628811. S2CID 17783758.
- ^ Donald McNeil, Jr. (September 16, 2010). "Precursor to H.I.V. was in monkeys for millennia". The New York Times. Retrieved September 17, 2010.
Dr. Marx believes that the crucial event was the introduction into Africa of millions of inexpensive, mass-produced syringes in the 1950s. ... suspect that the growth of colonial cities is to blame. Before 1910, no Central African town had more than 10,000 people. But urban migration rose, increasing sexual contacts and leading to red-light districts.
Further reading
- Berlier W, Bourlet T, Lawrence P, Hamzeh H, Lambert C, Genin C, Verrier B, Dieu-Nosjean MC, Pozzetto B, Delézay O (2005). "Selective sequestration of X4 isolates by human genital epithelial cells: Implication for virus tropism selection process during sexual transmission of HIV". Journal of Medical Virology. 77 (4): 465–74. doi:10.1002/jmv.20478. PMID 16254974. S2CID 25762969.
- Joint United Nations Programme on HIV/AIDS (UNAIDS) (2011). Global HIV/AIDS Response, Epidemic update and health sector progress towards universal access (PDF). Joint United Nations Programme on HIV/AIDS.
- Muciaccia B, Padula F, Vicini E, Gandini L, Lenzi A, Stefanini M (2005). "Beta-chemokine receptors 5 and 3 are expressed on the head region of human spermatozoon". The FASEB Journal. 19 (14): 2048–50. doi:10.1096/fj.05-3962fje. PMID 16174786. S2CID 7928126.
External links
- Definitions from Wiktionary
- Media from Wikimedia Commons
- News from Wikinews
- Textbooks from Wikibooks
- HIV/AIDS at Curlie