Клатрин - это белок, который играет важную роль в образовании везикул, покрытых оболочкой . Клатрин был впервые выделен и назван Барбарой Пирс в 1976 году. [1] Он образует форму трискелиона, состоящую из трех тяжелых цепей клатрина и трех легких цепей. Когда трискелии взаимодействуют, они образуют многогранную решетку, окружающую везикулу, отсюда и название белка, которое происходит от латинского clathrum, означающего решетку. Белки оболочки, такие как клатрин, используются для создания небольших пузырьков, чтобы транспортировать молекулы внутри клеток. Эндоцитоза и экзоцитозаВезикул позволяет клеткам общаться, передавать питательные вещества, импортировать сигнальные рецепторы, опосредовать иммунный ответ после отбора проб из внеклеточного мира и очищать клеточный мусор, оставленный воспалением ткани. Эндоцитарный путь может быть захвачен вирусами и другими патогенами, чтобы проникнуть в клетку во время инфекции. [2]
СОДЕРЖАНИЕ
1 Структура
2 Функция
3 См. Также
4 ссылки
5 Дальнейшее чтение
6 Внешние ссылки
Структура [ править ]
Клатрин легкая цепь а
Идентификаторы
Символ
CLTA
Ген NCBI
1211
HGNC
CLTA. HGNC: 2090. CLTA.
UniProt
P09496
Прочие данные
Locus
Chr. 9 q13
Ищи
Структуры
Швейцарская модель
Домены
ИнтерПро
Легкая цепь клатрина b
Идентификаторы
Символ
CLTB
Ген NCBI
1212
HGNC
2091
OMIM
118970
RefSeq
NM_001834
UniProt
P09497
Прочие данные
Locus
Chr. 5 q35
Ищи
Структуры
Швейцарская модель
Домены
ИнтерПро
Клатрин тяжелая цепь 1
Идентификаторы
Символ
CLTC
Альт. символы
CHC, CHC17, CLTCL2
Ген NCBI
1213
HGNC
2092
OMIM
118955
RefSeq
NM_004859
UniProt
Q00610
Прочие данные
Locus
Chr. 17 кв. 23,1 кв.
Ищи
Структуры
Швейцарская модель
Домены
ИнтерПро
Клатрин тяжелая цепь 2
Идентификаторы
Символ
CLTCL1
Альт. символы
CLTCL
Ген NCBI
8218
HGNC
2093
OMIM
601273
RefSeq
NM_001835
UniProt
P53675
Прочие данные
Locus
Chr. 22 q11.21
Ищи
Структуры
Швейцарская модель
Домены
ИнтерПро
Трискелион клатрина состоит из трех тяжелых цепей клатрина, взаимодействующих на своих С-концах , каждая тяжелая цепь массой ~ 190 кДа имеет прочно связанную с ней легкую цепь массой ~ 25 кДа. Три тяжелые цепи обеспечивают структурную основу решетки клатрина, а три легкие цепи, как полагают, регулируют образование и разборку решетки клатрина. Есть две формы легких цепей клатрина, обозначенные a и b. Основная тяжелая цепь клатрина, расположенная на хромосоме 17 у человека, обнаруживается во всех клетках. Второй ген тяжелой цепи клатрина на хромосоме 22 экспрессируется в мышцах.
Тяжелая цепь клатрина часто описывается как ножка с субдоменами, представляющими стопу ( N-концевой домен), за которой следуют лодыжка, дистальная часть ноги, колено, проксимальная часть ноги и домены тримеризации. N-концевой домен состоит из структуры β-пропеллера с семью лопастями. Остальные домены образуют суперспираль из коротких альфа-спиралей. Первоначально это было определено на основе структуры идентифицированного проксимального домена ноги, который состоит из меньшего структурного модуля, называемого мотивами повторов тяжелой цепи клатрина. Легкие цепи связываются в первую очередь с проксимальной ветвью тяжелой цепи с некоторым взаимодействием вблизи домена тримеризации. Β-пропеллер у «подножия» клатрина содержит множество сайтов связывания для взаимодействия с другими белками.
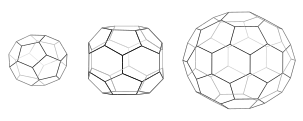
Каждая клетка состоит из 12 пятиугольников. Мини-покрытие (слева) имеет 4 шестиугольника и тетраэдрическую симметрию, как в усеченном трехугольном тетраэдре. Шестиугольный ствол (средний) имеет 8 шестиугольников и симметрию D6. Футбольный мяч (справа) имеет 20 шестиугольников и икосаэдрическую симметрию, как в усеченном икосаэдре.
Когда трискелии собираются вместе в растворе, они могут взаимодействовать с достаточной гибкостью, чтобы сформировать 6-сторонние кольца ( шестиугольники ), которые образуют плоскую решетку, или 5-сторонние кольца ( пятиугольники ), которые необходимы для образования криволинейной решетки. Когда много трискелионов соединяются, они могут образовывать структуру, похожую на корзину. Показанная структура состоит из 36 трискелий, одна из которых показана синим цветом. Еще одна распространенная сборка - усеченный икосаэдр . Чтобы заключить везикулу, в решетке должно присутствовать ровно 12 пятиугольников.
В клетке трискелион клатрина в цитоплазме связывается с адаптерным белком, который имеет связанную мембрану, соединяя один из своих трех футов с мембраной за раз. Клатрин не может напрямую связываться с мембраной или грузом и вместо этого использует для этого адаптерные белки. Этот трискелион будет связываться с другими прикрепленными к мембране трискелиями, образуя округлую решетку из шестиугольников и пятиугольников, напоминающую панели на футбольном мяче, которые вытягивают мембрану в бутон. Создавая различные комбинации 5-сторонних и 6-сторонних колец, везикулы разных размеров могут собираться. Самая маленькая клатриновая клетка, которую обычно изображают, называемая мини-пальто, имеет 12 пятиугольников и только два шестиугольника. Даже меньшие клетки с нулевыми шестиугольниками, вероятно, не образуются из природного белка, потому что ножки трискелий слишком громоздки.
Функция [ править ]
Механизм клатрин-опосредованного эндоцитоза.
Клатрин выполняет решающую роль в формировании округлых пузырьков в цитоплазме для внутриклеточного движения . Покрытые клатрином везикулы (CCV) выборочно сортируют груз на клеточной мембране , в сети транс- Гольджи и в эндосомных компартментах для множественных мембранных путей движения. После того, как везикула прорастает в цитоплазму, оболочка быстро разбирается, позволяя клатрину рециркулировать, в то время как везикула транспортируется в различные места.
Адаптерные молекулы ответственны за самосборку и набор. Двумя примерами адаптерных белков являются AP180 [3] и эпсин . [4] [5] [6] AP180 используется в формировании синаптических пузырьков. Он привлекает клатрин к мембранам, а также способствует его полимеризации.. Эпсин также привлекает клатрин к мембранам и способствует его полимеризации и может помочь деформировать мембрану, и, таким образом, везикулы, покрытые клатрином, могут образоваться. В клетке трискелион, плавающий в цитоплазме, связывается с адаптерным белком, соединяя одну из своих ножек с мембраной за раз. Скелион будет связываться с другими, прикрепленными к мембране, образуя многогранную решетку, скелион, которая вытягивает мембрану в бутон. Скелион не связывается напрямую с мембраной, но связывается с адапторными белками, которые распознают молекулы на поверхности мембраны.
Клатрин выполняет еще одну функцию, помимо покрытия органелл . В неделящихся клетках образование покрытых клатрином везикул происходит непрерывно. Формирование везикул, покрытых клатрином, прекращается в клетках, подвергающихся митозу . Во время митоза клатрин связывается с аппаратом веретена в комплексе с двумя другими белками: TACC3 и ch-TOG / CKAP5 . Клатрин помогает в конгрессе хромосом, стабилизируя кинетохорные волокна митотического веретена . Аминоконцевой домен тяжелой цепи клатрина и домен TACC TACC3 заставляют поверхность связывания микротрубочек для TACC3 / ch-TOG / клатрина связываться с митотическим веретеном.. Стабилизация кинетохорных волокон требует тримерной структуры клатрина для сшивания микротрубочек. [7] [8]
Клатрин-опосредованный эндоцитоз (CME) регулирует многие клеточные физиологические процессы, такие как интернализация факторов роста и рецепторов, проникновение патогенов и синаптическая передача. Считается, что захватчики клеток используют питательные пути, чтобы получить доступ к механизмам репликации клетки. Определенные сигнальные молекулы открывают путь питательных веществ. Два химических соединения, называемые Pitstop 1 и Pitstop 2, селективные ингибиторы клатрина, могут препятствовать патогенной активности и, таким образом, защищать клетки от вторжения. Эти два соединения избирательно блокируют ассоциацию эндоцитарного лиганда с концевым доменом клатрина in vitro. [9] Однако специфичность этих соединений для блокирования клатрин-опосредованного эндоцитоза подвергается сомнению. [10]
См. Также [ править ]
Адаптин
AP180
Формирование крышки
Миграция клеток
Dynamin
Эндоцитарный цикл
Эпсин
Синаптический пузырек
Ссылки [ править ]
^ Пирса BM (1976). «Клатрин: уникальный белок, связанный с внутриклеточным переносом мембраны покрытыми везикулами» . Труды Национальной академии наук Соединенных Штатов Америки . 73 (4): 1255–9. Bibcode : 1976PNAS ... 73.1255P . DOI : 10.1073 / pnas.73.4.1255 . PMC 430241 . PMID 1063406 .
^ "Архивная копия" . Архивировано 16 января 2016 года . Проверено 7 октября 2015 .CS1 maint: заархивированная копия как заголовок ( ссылка )
^ McMahon HT. «Клатрин и его взаимодействие с AP180» . Лаборатория молекулярной биологии MRC. Архивировано из оригинала на 2009-05-01 . Проверено 17 апреля 2009 . микрофотографии клатриновой сборки
^ McMahon HT. «Галерея Эпсин 1 ЭМ» . Лаборатория молекулярной биологии MRC. Архивировано из оригинала на 2009-01-02 . Проверено 17 апреля 2009 . микрофотографии почкования пузырьков
^ Форд MG, Пирс BM , Хиггинс MK, Валлис Y, Оуэн DJ, Gibson A, Хопкинс CR, Эванс PR, McMahon HT (февраль 2001). «Одновременное связывание PtdIns (4,5) P2 и клатрина AP180 при зарождении решеток клатрина на мембранах» (PDF) . Наука . 291 (5506): 1051–5. Bibcode : 2001Sci ... 291.1051F . CiteSeerX 10.1.1.407.6006 . DOI : 10.1126 / science.291.5506.1051 . PMID 11161218 . Архивировано из оригинального (PDF) 21 ноября 2008 года . Проверено 17 апреля 2009 .
Перейти ↑ Higgins MK, McMahon HT (2002). «Снимки клатрин-опосредованного эндоцитоза» (PDF) . Направления биохимических наук . 27 (5): 257–63. DOI : 10.1016 / S0968-0004 (02) 02089-3 . PMID 12076538 . Архивировано из оригинального (PDF) 21 ноября 2008 года . Проверено 17 апреля 2009 .
^ Royle SJ, Брайт Н.А., Lagnado L (2005). «Клатрин необходим для функции митотического веретена» . Природа . 434 (7037): 1152–1157. Bibcode : 2005Natur.434.1152R . DOI : 10,1038 / природа03502 . PMC 3492753 . PMID 15858577 .
^ Худ FE, Уильямс SJ, Берджесс SG, Ричардс MW, Roth D, Straube A, Pfuhl M, Bayliss R, Royle SJ (2013). «Координация соседних доменов опосредует сборку TACC3-ch-TOG-клатрин и связывание митотического веретена» . Журнал клеточной биологии . 202 (3): 463–78. DOI : 10,1083 / jcb.201211127 . PMC 3734082 . PMID 23918938 .
^ Роль терминального домена клатрины в Регулирующем ПОКРЫТИИ Pit Динамики Показала малую Молекулу Ингибирование | Cell, том 146, выпуск 3, 471-484, 5 августа 2011 Аннотация архивации 2012-01-19 в Wayback Machine
Фотин А., Ченг Ю., Слиз П., Григорьев Н., Харрисон С. К., Кирххаузен Т., Вальц Т. (2004). «Молекулярная модель полной решетки клатрина из электронной криомикроскопии». Природа . 432 (7017): 573–9. Bibcode : 2004Natur.432..573F . DOI : 10,1038 / природа03079 . PMID 15502812 . S2CID 4396282 .
Мусави С.А., Малед Л., Берг Т., Кьекен Р. (2004). «Клатрин-зависимый эндоцитоз» . Биохимический журнал . 377 (Pt 1): 1–16. DOI : 10.1042 / BJ20031000 . PMC 1223844 . PMID 14505490 .
Смит К.Дж., Григорьев Н., Пирс Б.М. (сентябрь 1998 г.). «Клатриновые оболочки с разрешением 21 A: клеточная сборка, предназначенная для рециркуляции множества мембранных рецепторов» . Журнал EMBO . 17 (17): 4943–53. DOI : 10.1093 / emboj / 17.17.4943 . PMC 1170823 . PMID 9724631 . (Модель сборки Clathrin)
Перес-Гомес Дж., Мур I. (март 2007 г.). «Эндоцитоз растений: все-таки клатрин». Текущая биология . 17 (6): R217–9. DOI : 10.1016 / j.cub.2007.01.045 . PMID 17371763 . S2CID 17680351 . (Обзор участия клатрина в эндоцитозе растений)
Ройл С.Дж., Брайт Н.А., Лагнадо Л. (апрель 2005 г.). «Клатрин необходим для функции митотического веретена» . Природа . 434 (7037): 1152–7. Bibcode : 2005Natur.434.1152R . DOI : 10,1038 / природа03502 . PMC 3492753 . PMID 15858577 .
Худ Ф. Э., Уильямс С. Дж., Берджесс С. Г., Ричардс М. В., Рот Д., Штрауб А., Пфул М., Бейлисс Р., Ройл С. Дж. (Август 2013 г.). «Координация соседних доменов опосредует сборку TACC3-ch-TOG-клатрин и связывание митотического веретена» . J Cell Biol . 202 (3): 463–78. DOI : 10,1083 / jcb.201211127 . PMC 3734082 . PMID 23918938 .
Knuehl C, Chen CY, Manalo V, Hwang PK, Ota N, Brodsky FM (2006). «Новые сайты связывания на клатрине и адаптерах регулируют отдельные аспекты сборки оболочки» . Трафик . 7 (12): 1688–700. DOI : 10.1111 / j.1600-0854.2006.00499.x . PMID 17052248 . S2CID 19087208 .
Эделинг М.А., Смит К., Оуэн Д. (2006). «Жизнь клатринового пальто: выводы из клатриновых и AP структур». Обзоры природы Молекулярная клеточная биология . 7 (1): 32–44. DOI : 10.1038 / nrm1786 . PMID 16493411 . S2CID 19393938 .


{kind=link}
{kind=link}