Эта статья требует дополнительных ссылок для проверки . ( март 2020 г. ) ( Узнайте, как и когда удалить это сообщение-шаблон ) |
{kind=link}
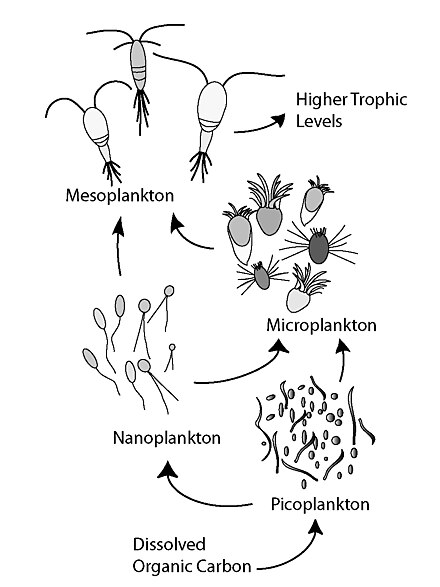
Микробной цикл описывает трофический путь в морской микробных пищевой сети , где растворенный органический углерод (DOC) возвращаются к более высоким трофическим уровням через его включение в бактериальную биомассу, а затем в сочетании с классической пищевой цепью , образованной фитопланктоном - зоопланктон - нектон . Термин микробная петля был введен Фаруком Азамом , Томом Фенчелом и др. [1] в 1983 г., чтобы включить роль бактерий в круговоротах углерода и питательных веществ в морской среде.
Как правило, растворенный органический углерод (DOC) попадает в океанскую среду в результате бактериального лизиса, утечки или экссудации фиксированного углерода из фитопланктона (например, слизистого экзополимера диатомовых водорослей ), внезапного клеточного старения, неаккуратного питания зоопланктоном, выделения отходов продукты от водных животных, или разложение или растворение органических частиц наземных растений и почв. [2] Бактерии в микробной петле разлагают этот детрит, чтобы использовать это богатое энергией вещество для роста. Поскольку более 95% органического вещества в морских экосистемах состоит из полимерных высокомолекулярных (ВММ) соединений (например, белков, полисахаридов, липидов), только небольшая часть общегорастворенное органическое вещество (РОВ) легко утилизируется большинством морских организмов на более высоких трофических уровнях. Это означает, что растворенный органический углерод недоступен для большинства морских организмов; морские бактерии вносят этот органический углерод в пищевую сеть, в результате чего дополнительная энергия становится доступной для более высоких трофических уровней. Недавно термин « микробная пищевая сеть » был заменен термином «микробная петля».
История [ править ]
| Часть серии о |
| Цикл углерода |
|---|
 |
До открытия микробной петли классический взгляд на морские пищевые сети представлял собой линейную цепочку от фитопланктона до нектона . Как правило, морские бактерии не считались значительными потребителями органических веществ (включая углерод), хотя было известно, что они существуют. Однако представление о морской пелагической пищевой сети было оспорено в 1970-х и 1980-х годах Померой и Азамом, которые предложили альтернативный путь потока углерода от бактерий к простейшим и многоклеточным . [3] [1]
Ранние работы в области морской экологии, которые исследовали роль бактерий в океанической среде, пришли к выводу, что их роль очень минимальна. Традиционные методы подсчета бактерий (например, культивирование на чашках с агаром ) давали лишь небольшое количество бактерий, которое было намного меньше их истинного количества в окружающей среде в морской воде. Развитие технологий для подсчета бактерий привело к пониманию важности морских бактерий в океанической среде.
В 1970-х годах Франциско и др. Разработали альтернативный метод прямого микроскопического счета . (1973) и Hobbie et al. (1977). Бактериальные клетки подсчитывали с помощью эпифлуоресцентного микроскопа , получая так называемый « прямой подсчет акридинового апельсина » (AODC). Это привело к переоценке большой концентрации бактерий в морской воде, которая оказалась больше, чем ожидалось (обычно порядка 1 миллиона на миллилитр). Кроме того, разработка «анализа бактериальной продуктивности» показала, что большая часть (т.е. 50%) чистой первичной продукции (NPP) была переработана морскими бактериями.
В 1974 году Ларри Помрой опубликовал в журнале BioScience статью под названием «Пищевая сеть океана: меняющаяся парадигма», в которой была подчеркнута ключевая роль микробов в продуктивности океана. [3] В начале 1980-х Азам и группа ведущих ученых-океанологов опубликовали в журнале « Marine Ecology Progress Series » обобщение своих дискуссий под названием «Экологическая роль микробов в водной толще в море». Термин «микробная петля» был введен в эту статью, в которой отмечается, что потребляющие бактерии простейшие принадлежат к тому же классу размеров, что и фитопланктон, и, вероятно, являются важным компонентом рациона планктонных ракообразных . [1]
Доказательства, накопленные с того времени, показали, что некоторые из этих бактериоядных простейших (например, инфузории ) на самом деле выборочно становятся жертвами этих веслоногих рачков . В 1986 году Салли В. Чизолм , Роберт Дж. Олсон и другие сотрудники обнаружили прохлорококк , который в большом количестве встречается в олиготрофных районах океана (хотя было несколько более ранних записей об очень мелких цианобактериях, содержащих хлорофилл b в океан [4] [5] Prochlorococcus был открыт в 1986 году [6] ). [7] Исходя из этого открытия, исследователи наблюдали изменение роли морских бактерий в градиенте питательных веществ от эвтрофных к олиготрофным областям океана.
Факторы, контролирующие микробный цикл [ править ]
Эффективность микробной петли определяется плотностью морских бактерий внутри нее. [8] Стало ясно, что плотность бактерий в основном контролируется пастбищной активностью мелких простейших и различных таксономических групп жгутиконосцев. Кроме того, вирусная инфекция вызывает бактериальный лизис, который высвобождает содержимое клеток обратно в растворенные органические вещества.(DOM) пул, снижая общую эффективность микробной петли. Смертность от вирусной инфекции почти такая же, как от выпаса простейших. Однако, по сравнению с выпасом простейших, эффект вирусного лизиса может сильно отличаться, потому что лизис сильно зависит от хозяина для каждой морской бактерии. Как выпас простейших, так и вирусная инфекция уравновешивают основную долю роста бактерий. Кроме того, микробная петля преобладает в олиготрофных водах, а не в эвтрофных районах - там преобладает классическая пищевая цепь планктона из-за частого поступления свежих минеральных питательных веществ (например, весеннее цветение в умеренных водах, районы апвеллинга ). Величину эффективности микробной петли можно определить путем измерения бактериального включения радиоактивно меченных субстратов (таких какмеченный тритием тимидин или лейцин).
Значение в морских экосистемах [ править ]
Микробный цикл имеет особое значение в повышении эффективности морской пищевой сети за счет использования растворенного органического вещества (РОВ), которое обычно недоступно для большинства морских организмов. В этом смысле процесс помогает рециркулировать органические вещества и питательные вещества и обеспечивает передачу энергии над термоклином . Более 30% растворенного органического углерода (DOC), содержащегося в бактериях, вдыхается и выделяется в виде диоксида углерода . Другой главный эффект микробной петли в толще воды заключается в том, что она ускоряет минерализацию.за счет восстановления производства в средах с ограниченными питательными веществами (например, в олиготрофных водах). В целом, вся микробная петля в некоторой степени обычно в пять-десять раз превышает массу всех многоклеточных морских организмов в морской экосистеме. Морские бактерии являются основой пищевой сети в большинстве океанических сред, и они повышают трофическую эффективность как морских пищевых сетей, так и важных водных процессов (таких как продуктивность рыболовства и количество углерода, экспортируемого на дно океана). Таким образом, микробный цикл вместе с первичной продукцией контролирует продуктивность морских систем в океане.
Многие планктонные бактерии подвижны, используют жгутик для размножения и хемотаксис, чтобы находить, перемещаться и прикрепляться к точечному источнику растворенного органического вещества (РОВ), где быстрорастущие клетки переваривают всю или часть частицы. Накопление в течение нескольких минут на таких участках непосредственно наблюдается. Следовательно, толщу воды можно рассматривать в некоторой степени как пространственно организованное место в небольшом масштабе, а не как полностью смешанную систему. Это образование пятен влияет на биологически опосредованный перенос вещества и энергии в микробной петле.
В настоящее время микробный цикл считается более протяженным. [9] Химические соединения в типичных бактериях (например, ДНК, липиды, сахара и т. Д.) И аналогичные значения соотношений C: N на частицу обнаруживаются в микрочастицах, образованных абиотически. Микрочастицы являются потенциально привлекательным источником пищи для бактериоядного планктона. В этом случае микробная петля может быть расширена путем прямого переноса растворенного органического вещества.(DOM) через образование абиотических микрочастиц на более высокие трофические уровни. Это имеет двоякое экологическое значение. Во-первых, это происходит без потери углерода и делает органическое вещество более доступным для фаготрофных организмов, а не только для гетеротрофных бактерий. Кроме того, абиотическая трансформация в расширенной микробной петле зависит только от температуры и способности РОВ к агрегации, в то время как биотическая трансформация зависит от его биологической доступности. [9]
См. Также [ править ]
- Биологический насос
- f-соотношение
- Планктон
- Морской снег
- Фикосфера
Ссылки [ править ]
- ^ a b c Азам, Фарук ; Фенчел, Том ; Филд, JG; Серый, JS; Meyer-Reil, LA; Тингстад, Ф. (1983). «Экологическая роль микробов водяного столба в море» . Серия «Прогресс морской экологии» . 10 : 257–263. DOI : 10,3354 / meps010257 .
- ^ Ван ден Меерш, Карел; Мидделбург, Джек Дж .; Soetaert, Karline; ван Рейсвейк, Питер; Бошкер, Хенрикус Т.С.; Хейп, Карло HR (2004). «Связь углерода и азота и взаимодействия водорослей и бактерий во время экспериментального цветения: моделирование эксперимента с трассером a13C». Лимнология и океанография . 49 (3): 862–878. DOI : 10,4319 / lo.2004.49.3.0862 . ISSN 0024-3590 .
- ^ a b Помрой, Лоуренс Р. (1974). «Пищевая сеть океана, меняющаяся парадигма». Бионаука . 24 (9): 499–504. DOI : 10.2307 / 1296885 . ISSN 0006-3568 .
- ^ Джонсон, PW; Sieburth, JM (1979). «Хроококковые цианобактерии в море: повсеместная и разнообразная фототрофная биомасса». Лимнология и океанография . 24 (5): 928–935. Bibcode : 1979LimOc..24..928J . DOI : 10,4319 / lo.1979.24.5.0928 .
- ^ Gieskes, WWC; Краай, GW (1983). «Неизвестные хлорофилл а производные в Северном море и в тропической части Атлантического океана показали с помощью анализа ВЭЖХ» . Лимнология и океанография . 28 (4): 757–766. Bibcode : 1983LimOc..28..757G . DOI : 10,4319 / lo.1983.28.4.0757 .
- ^ Chisholm, SW; Олсон, Р.Дж.; Зеттлер, ER; Waterbury, J .; Goericke, R .; Вельшмайер, Н. (1988). «Новый свободноживущий прохлорофит встречается при высоких концентрациях клеток в океанической эвфотической зоне». Природа . 334 (6180): 340–343. Bibcode : 1988Natur.334..340C . DOI : 10.1038 / 334340a0 .
- ^ Чисхолм, Салли В .; Франкель, Шейла Л .; Герике, Ральф; Олсон, Роберт Дж .; Паленик, Брайан; Уотербери, Джон Б.; Вест-Джонсруд, Лиза; Зеттлер, Эрик Р. (1992). «Prochlorococcus marinus nov. Gen. Nov. Sp .: оксифотрофный морской прокариот, содержащий дивинилхлорофиллы a и b». Архив микробиологии . 157 (3): 297–300. DOI : 10.1007 / bf00245165 . ISSN 0302-8933 .
- ^ Тейлор, AH; Джойнт, I (1990). «Стационарный анализ« микробной петли »в стратифицированных системах» . Серия «Прогресс морской экологии» . Межисследовательский научный центр. 59 : 1–17. DOI : 10,3354 / meps059001 . ISSN 0171-8630 .
- ^ а б Кернер, Мартин; Хоэнберг, Хайнц; Эртль, Зигмунд; Рекерманн, Маркус; Спитзи, Алехандро (2003). «Самоорганизация растворенного органического вещества в мицеллоподобные микрочастицы в речной воде». Природа . ООО "Спрингер Сайенс энд Бизнес Медиа". 422 (6928): 150–154. DOI : 10,1038 / природа01469 . ISSN 0028-0836 .
Библиография [ править ]
- Фенчел, Т. (1988) Морские планктонные пищевые цепи. Ежегодный обзор экологии и систематики
- Фенчел, Т. (2008) Микробная петля - 25 лет спустя. Журнал экспериментальной морской биологии и экологии
- Fuhrman, JA, Azam, F. (1982) Включение тимидина как мера продукции гетеротрофного бактериопланктона в морских поверхностных водах. Морская биология
- Кернер, М., Хоэнберг, Х., Эртл, С., Рекерманн, М., Спитзи, А. (2003) Самоорганизация растворенного органического вещества в мицеллоподобные микрочастицы в речной воде. Природа
- Кирчман Д., Сигда Дж., Капусцински Р., Митчелл Р. (1982) Статистический анализ метода прямого подсчета для подсчета бактерий. Прикладная и экологическая микробиология
- Мейнхард С., Азам Ф. (1989) Содержание белка и скорость синтеза белка планктонными морскими бактериями. Серия "Прогресс морской экологии"
- Мюнстер, В.Ю. (1985) Исследования структуры, распределения и динамики различных органических субстратов в РОВ озера Плюссзее. Гидробиология
- Помрой, Л. Р., Уильямс, П. Дж. Б., Азам, Ф. и Хобби, Д. Е. (2007) "Микробная петля". Океанография , 20 (2): 28–33. DOI : 10,4319 / lo.2004.49.3.0862 .
- Стодереггер, К., Херндл, Дж. Дж. (1998) Производство и высвобождение бактериального капсульного материала и его последующее использование морским бактериопланктоном. Лимнология и океанография