| Рибозим вируса гепатита дельта | |
|---|---|
 Прогнозируемая вторичная структура и сохранение последовательности рибозима HDV | |
| Идентификаторы | |
| Символ | HDV_ribozyme |
| Рфам | RF00094 |
| Прочие данные | |
| Тип РНК | Джин ; рибозим |
| Домен (ы) | Вирусы |
| ТАК | ТАК: 0000374 |
| Структуры PDB | PDBe |
{kind=link}
{kind=link}
Вируса гепатита дельта (HDV) рибозят является некодирующей РНК найдена в вирусе гепатита дельты , который необходим для репликации вируса и является единственным известным человеком вирусом , который использует рибозима активность инфицировать свой хозяин. [1] рибозим действует , чтобы обрабатывать РНК - транскриптов в длину блок в саморасщепления реакции в ходе репликации вируса гепатита дельта, который , как полагают, распространяются двойным механизмом прокатки круга. [2] [3] Рибозим активен in vivo в отсутствие каких-либо белковых факторов и на момент своего открытия был самой быстрой из известных естественных саморасщепляющихся РНК. [4]
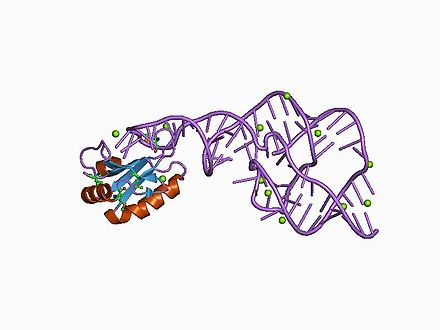
Кристаллическая структура этого рибозима была решена с помощью рентгеновской кристаллографии и показывает пять спиральных сегментов, соединенных двойным псевдоузлом . [1]
В дополнение к смысловой (геномной версии) все вирусы HDV также имеют антигеномную версию рибозима HDV. [5] Эта версия не является точной комплементарной последовательностью, но имеет ту же структуру, что и смысловая (геномная) цепь. Единственные «существенные» различия между ними - это небольшая выпуклость на стержне P4 и более короткое соединение J4 / 2. Для репликации необходимы как геномные, так и антигеномные рибозимы. [2]
HDV-подобные рибозимы [ править ]
Рибозим HDV структурно и биохимически родственен многим другим саморасщепляющимся рибозимам. Эти другие рибозимы часто называют примерами рибозимов HDV из-за их сходства, хотя они не обнаруживаются в вирусах гепатита дельта. Их также можно назвать «подобными HDV», чтобы указать на этот факт. [6]
HDV-подобные рибозимы включают рибозим CPEB3 млекопитающих , члены ретротранспозонов (например, в элементе РНК R2 у насекомых и в L1Tc и, возможно, в других ретротранспозонах у трипаносоматид) и последовательности из бактерий. [7] [8] [6] [9] [10] Группировка, вероятно, является результатом конвергентной эволюции : дельтавирус, обнаруженный за пределами человека, также обладает рибозимом DV, и ни один из предложенных сценариев горизонтального переноса генов еще не может объяснить это. [11] [12]
Механизм катализа [ править ]
Рибозим HDV катализирует разрыв фосфодиэфирной связи между нуклеотидом или олигонуклеотидом субстрата и 5'-гидроксилом рибозима. В вирусе гепатита дельта эта нуклеотидная последовательность субстрата начинается с уридина и известна как U (-1), однако идентичность нуклеотида -1 существенно не меняет скорость катализа. [1] Требуется только его химическая природа, поскольку, как показали Перротта и Бин, замещение U (-1) рибозы дезоксирибозой отменяет реакцию, что согласуется с предсказанием, что 2'-гидроксил является нуклеофил в химической реакции. [13]Следовательно, в отличие от многих других рибозимов, таких как рибозим «головка молотка», рибозим HDV не имеет дополнительных требований для катализа и для эффективной реакции требуется только один -1-рибонуклеотид в качестве субстрата. [1]
Первоначально считалось, что 75-й нуклеотид в рибозиме, цитозин, известный как C75, способен действовать как общее основание с N3 C75, отщепляющим протон от 2'-гидроксила U (-1) нуклеотида до облегчают нуклеофильную атаку на фосфодиэфирную связь. [1] Однако, хотя хорошо известно, что N3 C75 имеет pKa, возмущенный от его нормального значения 4,45 и ближе к примерно 6,15 или 6,40, [14] [15] он недостаточно нейтрален, чтобы действовать как общее базовый катализатор. Вместо этого N3 в C75, как полагают, действует как кислота Льюиса, стабилизируя уходящий 5'-гидроксил рибозима; это подтверждается его близостью к 5'-гидроксилу в кристаллической структуре. [1] [16]Замена нуклеотида C75 любым другим нуклеотидом отменяет или существенно ухудшает активность рибозима, хотя эта активность может быть частично восстановлена имидазолом, что дополнительно влияет на каталитическую активность C75. [17]
C75 в рибозиме HDV был предметом нескольких исследований из-за его специфической pKa. Типичные значения pKa для свободных нуклеозидов составляют от 3,5 до 4,2; эти более низкие значения pKa являются кислыми, и маловероятно, что они станут основными. Однако вполне вероятно, что структурная среда внутри рибозима, которая включает десольватированную щель активного сайта, обеспечивает отрицательный электростатический потенциал, который может возмущать pKa цитозина в достаточной степени, чтобы действовать как кислота Льюиса. [18] [19] [20] [21]
В дополнение к стабилизации кислотой Льюиса уходящей 5'-гидроксильной группы, теперь также принято, что рибозим HDV может использовать ион металла, чтобы способствовать активации 2'-гидроксила для атаки на U (-1) нуклеотид. Ион магния в активном центре рибозима координирован с 2'-гидроксильным нуклеофилом и кислородом ножничного фосфата и может действовать как кислота Льюиса для активации 2'-гидроксила. [17] [16] Кроме того, возможно, что фосфат U23 может действовать как кислота Льюиса, принимая протон от 2'-гидроксила с магнием, выступающим в качестве координирующего иона. [22]Поскольку рибозим HDV не требует активности ионов металлов, он не является облигатным металлоферментом, но присутствие магния в активном центре значительно улучшает реакцию расщепления. Рибозим HDV, по-видимому, имеет неспецифическую потребность в небольших количествах двухвалентных катионов для сворачивания, будучи активным в Mg 2+ , Ca 2+ , Mn 2+ и Sr 2+ . [1] В отсутствие ионов металлов кажется вероятным, что вода может заменить магний в качестве кислоты Льюиса.
Регулирование вышестоящей РНК [ править ]
В этом разделе отсутствует информация об иллюстрации различных состояний SS (см. PMID 17956974 , необходимо перерисовать с помощью R2R). ( Май 2020 г. ) |
Из-за того, что рибозим HDV является быстрым саморасщеплением, предыдущие эксперименты с рибонуклеазой проводили на 3'-продукте саморасщепления, а не на его предшественнике. [23] Однако известно, что фланкирующая последовательность участвует в регуляции активности саморасщепления рибозима HDV. [24] [25] [26] Таким образом, для изучения результирующей активности саморасщепления рибозима HDV была включена предшествующая последовательность 5 'от сайта саморасщепления. [24] Были определены две альтернативные структуры.
Первая ингибирующая структура складывается с помощью расширенного транскрипта (то есть транскрипта -30/99, координаты указаны относительно сайта саморасщепления), охватывающего от 30 нуклеотидов перед сайтом расщепления до 15 нуклеотидов ниже 3'-конца. [24] Фланкирующая последовательность изолирует рибозим в кинетической ловушке во время транскрипции и приводит к чрезвычайно низкой скорости саморасщепления. [24] Эта предотвращающая саморасщепление структура включает 3 альтернативных основы: Alt1, Alt2 и Alt3, которые нарушают активную конформацию. Alt1 представляет собой дальнодействующее взаимодействие длиной 10 пар оснований, образованное ингибирующим участком выше по течению (-25 / -15 нуклеотидов) и ниже по течению (76/86 нуклеотидов). [24]Alt1 разрушает стебель P2 в активной конформации, при этом предполагается, что P2 играет активирующую роль как для геномного, так и для антигеномного рибозима. [24] [27] [28] Alt2 представляет собой взаимодействие между вышестоящей фланкирующей последовательностью и рибозимом, а Alt3 представляет собой ненативное взаимодействие рибозим-рибозим. [24]
Вторичная структура этой ингибирующей конформации подтверждается различными экспериментальными подходами. [24] Во-первых, было выполнено прямое зондирование с помощью рибонуклеаз, а последующее моделирование с помощью mfold 3.0 с использованием ограничений из результатов зондирования согласуется с предложенной структурой. [24] Во-вторых, ряд олигомеров ДНК, комплементарных различным областям AS1 / 2, был использован для восстановления активности рибозима; результаты подтверждают ингибирующую роль AS1 / 2. [24] В- третьих, мутационный анализ вводит одиночные / двойные мутации вне рибозима, чтобы гарантировать, что наблюдаемая активность рибозима напрямую связана со стабильностью Alt1. [24] Было обнаружено, что стабильность AS1 обратно пропорциональна активности саморасщепления.[24]
Вторая пермиссивная структура позволяет рибозиму HDV саморасщепляться котранскрипционно, и эта структура дополнительно включает часть -54 / -18 нуклеотидов транскрипта РНК. [24] Находящийся выше ингибирующий участок -24 / -15 от вышеупомянутой ингибирующей конформации теперь изолирован в шпильке P (-1), расположенной выше сайта расщепления. [24] [29] [30] Мотив P (-1), однако, обнаруживается только в геномной последовательности, что может быть коррелировано с явлением, что геномные копии РНК HDV более многочисленны в инфицированных клетках печени. [24] [5]Экспериментальные данные также подтверждают эту альтернативную структуру. Во-первых, структурное картирование с помощью рибонуклеазы используется для зондирования фрагмента -54 / -1 вместо всего транскрипта-предшественника из-за быстро расщепляющейся природы этой структуры, которая согласуется с локальной шпилькой P (-1) (между -54 / -40 и -18 / -30 нт). [24] Во-вторых, эволюционная консервация обнаружена в P (-1) и связывающей области между P (-1) и P1 среди 21 геномного изолята РНК HDV. [24]
Использование в подготовке транскриптов РНК [ править ]
Особые свойства реакции расщепления рибозима HDV делают его полезным инструментом для получения транскриптов РНК с гомогенными 3'-концами, альтернативой транскрипции РНК с помощью РНК-полимеразы Т7, которая часто может давать гетерогенные концы или нежелательные добавления. Вариант кДНК рибозима может быть получен рядом с кДНК последовательности РНК-мишени и РНК, полученной путем транскрипции с помощью РНК-полимеразы Т7. Последовательность рибозима будет эффективно отщепляться без дополнительных требований, поскольку -1 нуклеотид является инвариантным, оставляя 2'– 3 'циклический фосфат, который можно легко удалить обработкой фосфатазой или полинуклеотидкиназой Т4. [31] Затем целевую РНК можно очистить с помощью гель-очистки.
Ссылки [ править ]
- ^ a b c d e f g h Ферре-Д'Амаре А. Р., Чжоу К., Дудна Дж. А. (октябрь 1998 г.). «Кристаллическая структура рибозима вируса гепатита дельта». Природа . 395 (6702): 567–574. Bibcode : 1998Natur.395..567F . DOI : 10.1038 / 26912 . PMID 9783582 .
- ^ a b Modahl LE, Lai MM (июль 1998 г.). «Транскрипция мРНК антигена гепатита дельта продолжается на протяжении всей репликации вируса гепатита дельта (HDV): новая модель транскрипции и репликации РНК HDV» . Журнал вирусологии . 72 (7): 5449–5456. PMC 110180 . PMID 9621000 .
- ^ Макнотон TB, Ши ST, Modahl LE, Lai MM (апрель 2002). «Репликация РНК вируса гепатита дельта по катящемуся кругу осуществляется двумя различными клеточными РНК-полимеразами» . Журнал вирусологии . 76 (8): 3920–3927. DOI : 10,1128 / JVI.76.8.3920-3927.2002 . PMC 136092 . PMID 11907231 .
- ↑ Kuo MY, Sharmeen L, Dinter-Gottlieb G, Taylor J (декабрь 1988 г.). «Характеристика последовательностей саморасщепляющейся РНК на геноме и антигеноме дельта-вируса гепатита человека» . Журнал вирусологии . 62 (12): 4439–4444. PMC 253552 . PMID 3184270 .
- ^ a b Чен П.Дж., Калпана Дж., Голдберг Дж., Мейсон В., Вернер Б., Герин Дж., Тейлор Дж. (ноябрь 1986 г.). «Структура и репликация генома вируса гепатита дельта» . Труды Национальной академии наук Соединенных Штатов Америки . 83 (22): 8774–8778. DOI : 10.1073 / pnas.83.22.8774 . PMC 387014 . PMID 2430299 .
- ^ a b Webb CH, Lupták A (2011). «HDV-подобные саморасщепляющиеся рибозимы» . Биология РНК . 8 (5): 719–727. DOI : 10,4161 / rna.8.5.16226 . PMC 3256349 . PMID 21734469 .
- ^ Eickbush DG, Eickbush TH (июль 2010). «Ретротранспозоны R2 кодируют саморасщепляющийся рибозим для процессинга из котранскрипта рРНК» . Молекулярная и клеточная биология . 30 (13): 3142–3150. DOI : 10.1128 / MCB.00300-10 . PMC 2897577 . PMID 20421411 .
- ^ Webb CH, Riccitelli NJ, Ruminski DJ, Lupták A (ноябрь 2009). «Широкое распространение саморасщепляющихся рибозимов» . Наука . 326 (5955): 953. Bibcode : 2009Sci ... 326..953W . DOI : 10.1126 / science.1178084 . PMC 3159031 . PMID 19965505 .
- ^ Санчес-Лука FJ, Лопез MC, Масиас F, Алонсо C, Томас MC (октябрь 2011). «Идентификация рибозима, подобного вирусу гепатита дельта, на 5'-конце мРНК ретротранспозона L1Tc из Trypanosoma cruzi» . Исследования нуклеиновых кислот . 39 (18): 8065–8077. DOI : 10.1093 / NAR / gkr478 . PMC 3185411 . PMID 21724615 .
- ^ Санчес-Лука F, Лопез MC, Масиас F, Алонсо C, Томас MC (январь 2012). «Pr77 и L1TcRz: двойная система на 5'-конце ретротранспозона L1Tc, внутренний промотор и HDV-подобный рибозим» . Мобильные генетические элементы . 2 (1): 1–7. DOI : 10.4161 / mge.19233 . PMC 3383444 . PMID 22754746 .
- ^ Hetzel U, Szirovicza L, Smura T, Prähauser B, Vapalahti O, Kipar A, Hepojoki J (апрель 2019). «Идентификация нового дельтавируса у удавов» . mBio . 10 (2). DOI : 10,1128 / mBio.00014-19 . PMC 6445931 . PMID 30940697 .
- ^ Chang WS, Петерсон JH, Le Lay C, Ши M, Lo N, M Wille, Eden JS, Holmes EC (июль 2019). «Новые агенты, подобные гепатиту D, у позвоночных и беспозвоночных» . Эволюция вирусов . 5 (2): vez021. DOI : 10,1093 / ве / vez021 . PMC 6628682 . PMID 31321078 .
- ^ Перротта, AT; Бин, Мэриленд (14 января 1992 г.). «Расщепление олигорибонуклеотидов рибозимом, полученным из последовательности РНК вируса гепатита дельта». Биохимия . 31 (1): 16–21. DOI : 10.1021 / bi00116a004 . PMID 1731868 .
- ^ Ли TS, Радак BK, Harris ME, York DM (2016). «Путь конформационного переключения с двумя металлическими ионами для активации рибозима HDV» . Катализ ACS . 6 (3): 1853–1869. DOI : 10.1021 / acscatal.5b02158 . PMC 5072530 . PMID 27774349 .
- ^ Gong B, Chen JH, Chase E, Chadalavada DM, Ядзима R, Golden BL, Bevilacqua PC, Кэри PR (октябрь 2007). «Прямое измерение рК (а), близкого к нейтральному, для каталитического цитозина в геномном рибозиме HDV с использованием рамановской кристаллографии». Журнал Американского химического общества . 129 (43): 13335–13342. DOI : 10.1021 / ja0743893 . PMID 17924627 .
- ^ а б Чен Дж. Х., Ядзима Р., Чадалавада Д. М., Чейз Е., Bevilacqua PC, Golden BL (август 2010 г.). «Кристаллическая структура 1,9 предварительного расщепления рибозима HDV предполагает, что как кислота Льюиса, так и общие кислотные механизмы вносят вклад в расщепление фосфодиэфира». Биохимия . 49 (31): 6508–6518. DOI : 10.1021 / bi100670p . PMID 20677830 .
- ^ a b Накано С., Проктор DJ, Bevilacqua PC (октябрь 2001 г.). «Механистическая характеристика геномного рибозима HDV: оценка каталитического и структурного вклада ионов двухвалентных металлов в механизм многоканальной реакции». Биохимия . 40 (40): 12022–12038. DOI : 10.1021 / bi011253n . PMID 11580278 .
- ^ Rajagopal P, Feigon J (июнь 1989). «Образование тройной цепи в гомопурине: гомопиримидиновые ДНК-олигонуклеотиды d (GA) 4 и d (TC) 4». Природа . 339 (6226): 637–640. DOI : 10.1038 / 339637a0 . PMID 2733796 .
- ^ Sklenár В, Feigon J (июнь 1990 г.). «Формирование стабильного триплекса из одной нити ДНК». Природа . 345 (6278): 836–838. Bibcode : 1990Natur.345..836S . DOI : 10.1038 / 345836a0 . PMID 2359461 .
- ^ Коннелл GJ, Ярус M (май 1994). «РНК с двойной специфичностью и двойные РНК со сходной специфичностью». Наука . 264 (5162): 1137–1141. Bibcode : 1994Sci ... 264.1137C . DOI : 10.1126 / science.7513905 . PMID 7513905 .
- ^ Лего P, Парди A (сентябрь 1994). «Исследование протонирования аденина в РНК на месте с помощью 13C ЯМР». Журнал Американского химического общества . 116 (18): 8390–8391. DOI : 10.1021 / ja00097a066 .
- ^ Каспрович А, Kempińska А, Smólska В, Wrzesiński Дж, Ciesiołka J (2015). «Применение флуоресцентно меченного транс-действующего антигеномного рибозима HDV для мониторинга взаимодействий антибиотик-РНК». Аналитические методы . 7 (24): 10414–10421. DOI : 10.1039 / C5AY02953H .
- Перейти ↑ Rosenstein SP, Been MD (октябрь 1991 г.). «Доказательства того, что элементы саморасщепляющейся геномной и антигеномной РНК вируса гепатита дельта имеют сходные вторичные структуры» . Исследования нуклеиновых кислот . 19 (19): 5409–5416. DOI : 10.1093 / NAR / 19.19.5409 . PMC 328906 . PMID 1923826 .
- ^ a b c d e f g h i j k l m n o p q Чадалавада Д. М., Кнудсен С. М., Накано С., Bevilacqua PC (август 2000 г.). «Роль вышестоящей структуры РНК в облегчении каталитической складки рибозима геномного дельта-вируса гепатита». Журнал молекулярной биологии . 301 (2): 349–367. DOI : 10.1006 / jmbi.2000.3953 . PMID 10926514 .
- ^ Perrotta AT, Been MD (декабрь 1990). «Саморасщепляющийся домен геномной РНК вируса гепатита дельта: требования к последовательности и эффекты денатурирующего средства» . Исследования нуклеиновых кислот . 18 (23): 6821–6827. DOI : 10.1093 / NAR / 18.23.6821 . PMC 332737 . PMID 2263447 .
- ^ Perrotta AT, Been MD (апрель 1991). «Псевдоузелеподобная структура, необходимая для эффективного самоотщепления РНК вируса гепатита дельта». Природа . 350 (6317): 434–436. Bibcode : 1991Natur.350..434P . DOI : 10.1038 / 350434a0 . PMID 2011192 .
- ^ Matysiak M, Wrzesinski J, Ciesiołka J (август 1999). «Последовательная укладка геномного рибозима вируса гепатита дельта: структурный анализ промежуточных продуктов транскрипции РНК». Журнал молекулярной биологии . 291 (2): 283–294. DOI : 10.1006 / jmbi.1999.2955 . PMID 10438621 .
- ^ Perrotta AT, Никифорова O, Been MD (февраль 1999). «Консервативный выпуклый аденозин в периферическом дуплексе антигеномной саморасщепляющейся РНК HDV снижает кинетический захват неактивных конформаций» . Исследования нуклеиновых кислот . 27 (3): 795–802. DOI : 10.1093 / NAR / 27.3.795 . PMC 148249 . PMID 9889275 .
- ^ Mathews DH, Сабина J, Цукер M, Turner DH (Май 1999). «Расширенная зависимость термодинамических параметров от последовательности улучшает предсказание вторичной структуры РНК». Журнал молекулярной биологии . 288 (5): 911–940. DOI : 10.1006 / jmbi.1999.2700 . PMID 10329189 .
- ^ Цукер M, Mathews D, D Turner (1999). «Практическое руководство по алгоритмам и термодинамике для предсказания вторичной структуры РНК». В Кларке JB (ред.). Биохимия и биотехнология РНК . Серия НАТО ASI. Дордрехт, Нидерланды: Kluwer Academic Publishers.
- ^ Wichlacz A, Legiewicz M, Ciesiołka J (февраль 2004). «Создание транскриптов in vitro с гомогенными 3'-концами с использованием транс-действующего антигеномного дельта-рибозима» . Исследования нуклеиновых кислот . 32 (3): 39e – 39. DOI : 10.1093 / NAR / gnh037 . PMC 373431 . PMID 14973333 .
Внешние ссылки [ править ]
- Страница для рибозима вируса гепатита дельта в Rfam
- Запись в базе данных субвирусной РНК для рибозима HDV