| Рибозим вируса гепатита дельта | |
|---|---|
 Прогнозируемая вторичная структура и сохранение последовательности рибозима HDV | |
| Идентификаторы | |
| Символ | HDV_ribozyme |
| Рфам | RF00094 |
| Прочие данные | |
| Тип РНК | Джин ; рибозим |
| Домен (ы) | Вирусы |
| ТАК | ТАК: 0000374 |
| Структуры PDB | PDBe |
{kind=link}
{kind=link}
Вируса гепатита дельта (HDV) рибозят является некодирующей РНК найдена в вирусе гепатита дельты , который необходим для репликации вируса и является единственным известным человеком вирусом , который использует рибозима активность инфицировать свой хозяин. [1] рибозим действует , чтобы обрабатывать РНК - транскриптов в длину блок в саморасщепления реакции в ходе репликации вируса гепатита дельта, который , как полагают, распространяются двойным механизмом прокатки круга. [2] [3] Рибозим активен in vivo в отсутствие каких-либо белковых факторов и на момент своего открытия был самой быстрой из известных естественных саморасщепляющихся РНК. [4]
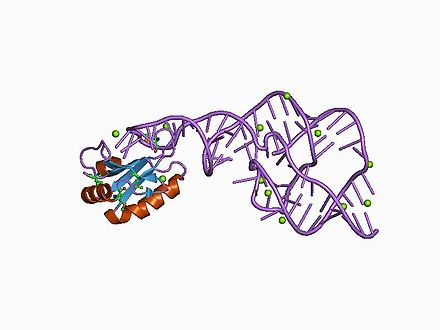
Кристаллическая структура этого рибозима была решена с помощью рентгеновской кристаллографии и показывает пять спиральных сегментов, соединенных двойным псевдоузлом . [1]
В дополнение к смысловой (геномной версии) все вирусы HDV также имеют антигеномную версию рибозима HDV. [5] Эта версия не является точной комплементарной последовательностью, но имеет ту же структуру, что и смысловая (геномная) цепь. Единственные «существенные» различия между ними - это небольшая выпуклость на стержне P4 и более короткое соединение J4 / 2. Для репликации необходимы как геномные, так и антигеномные рибозимы. [2]
HDV-подобные рибозимы [ править ]
Рибозим HDV структурно и биохимически родственен многим другим саморасщепляющимся рибозимам. Эти другие рибозимы часто называют примерами рибозимов HDV из-за их сходства, даже если они не обнаруживаются в вирусах гепатита дельта. Их также можно назвать «подобными HDV», чтобы указать на этот факт. [6]
HDV-подобные рибозимы включают рибозим CPEB3 млекопитающих , члены ретротранспозонов (например, в элементе РНК R2 у насекомых и в L1Tc и, возможно, в других ретротранспозонах у трипаносоматид) и последовательности из бактерий. [7] [8] [6] [9] [10] Группировка, вероятно, является результатом конвергентной эволюции : дельтавирус, обнаруженный за пределами человека, также обладает рибозимом DV, и ни один из предложенных сценариев горизонтального переноса генов еще не может объяснить это. [11] [12]
Механизм катализа [ править ]
Рибозим HDV катализирует расщепление фосфодиэфирной связи между нуклеотидом или олигонуклеотидом субстрата и 5'-гидроксилом рибозима. В вирусе гепатита дельта эта нуклеотидная последовательность субстрата начинается с уридина и известна как U (-1), однако идентичность нуклеотида -1 существенно не меняет скорость катализа. [1] Требуется только его химическая природа, поскольку, как показали Перротта и Бин, замещение U (-1) рибозы дезоксирибозой отменяет реакцию, что согласуется с предсказанием о том, что 2'-гидроксил является нуклеофил в химической реакции. [13]Следовательно, в отличие от многих других рибозимов, таких как рибозим «голова молотка», рибозим HDV не имеет дополнительных требований для катализа и требует только одного -1-рибонуклеотида в качестве субстрата для эффективной реакции. [1]
Первоначально считалось, что 75-й нуклеотид в рибозиме, цитозин, известный как C75, может действовать как общее основание с N3 C75, отщепляя протон от 2'-гидроксила U (-1) нуклеотида до облегчают нуклеофильную атаку на фосфодиэфирную связь. [1] Однако, хотя хорошо известно, что N3 C75 имеет pKa, возмущенное от его нормального значения 4,45 и ближе к примерно 6,15 или 6,40, [14] [15] оно недостаточно нейтрально, чтобы действовать как общее базовый катализатор. Вместо этого предполагается, что N3 в C75 действует как кислота Льюиса, стабилизируя уходящий 5'-гидроксил рибозима; это подтверждается его близостью к 5'-гидроксилу в кристаллической структуре. [1] [16]Замена нуклеотида C75 на любой другой нуклеотид отменяет или существенно снижает активность рибозима, хотя эта активность может быть частично восстановлена имидазолом, что дополнительно влияет на каталитическую активность C75. [17]
C75 в рибозиме HDV был предметом нескольких исследований из-за его специфической pKa. Типичные значения pKa для свободных нуклеозидов составляют от 3,5 до 4,2; эти более низкие значения pKa являются кислыми, и маловероятно, что они станут основными. Однако вполне вероятно, что структурная среда внутри рибозима, которая включает десольватированную щель активного сайта, обеспечивает отрицательный электростатический потенциал, который может возмущать pKa цитозина, достаточный для того, чтобы действовать как кислота Льюиса. [18] [19] [20] [21]
В дополнение к стабилизации кислотой Льюиса уходящей 5'-гидроксильной группы, теперь также считается, что рибозим HDV может использовать ион металла, чтобы способствовать активации 2'-гидроксила для атаки на U (-1) нуклеотид. Ион магния в активном центре рибозима координирован с 2'-гидроксильным нуклеофилом и кислородом ножничного фосфата и может действовать как кислота Льюиса для активации 2'-гидроксила. [17] [16] Кроме того, возможно, что фосфат U23 может действовать как кислота Льюиса, принимая протон от 2'-гидроксила с магнием, выступающим в качестве координирующего иона. [22]Поскольку рибозим HDV не требует активности ионов металлов, он не является облигатным металлоферментом, но присутствие магния в активном центре значительно улучшает реакцию расщепления. Рибозим HDV, по-видимому, имеет неспецифическую потребность в небольших количествах двухвалентных катионов для сворачивания, будучи активным в Mg 2+ , Ca 2+ , Mn 2+ и Sr 2+ . [1] В отсутствие ионов металлов кажется вероятным, что вода может заменить магний в качестве кислоты Льюиса.
Регулирование вышестоящей РНК [ править ]
В этом разделе отсутствует информация об иллюстрации различных состояний SS (см. PMID 17956974 , необходимо перерисовать с помощью R2R) . ( Май 2020 г. ) |
Из-за того, что рибозим HDV является быстрым саморасщеплением, предыдущие эксперименты с рибонуклеазой проводились с 3'-продуктом самоотщепления, а не с предшественником. [23] Однако известно, что фланкирующая последовательность участвует в регуляции активности саморасщепления рибозима HDV. [24] [25] [26] Таким образом, для изучения результирующей активности саморасщепления рибозима HDV была включена предшествующая последовательность 5 'от сайта саморасщепления. [24] Были определены две альтернативные структуры.
Первая ингибирующая структура складывается с помощью расширенного транскрипта (то есть транскрипта -30/99, координаты указаны относительно сайта саморасщепления), охватывающего от 30 нуклеотидов перед сайтом расщепления до 15 нуклеотидов ниже 3'-конца. [24] Фланкирующая последовательность изолирует рибозим в кинетической ловушке во время транскрипции и приводит к чрезвычайно низкой скорости саморасщепления. [24] Эта предотвращающая саморасщепление структура включает 3 альтернативных основы: Alt1, Alt2 и Alt3, которые нарушают активную конформацию. Alt1 представляет собой дальнодействующее взаимодействие длиной 10 пар оснований, образованное ингибирующим участком выше по течению (-25 / -15 нуклеотидов) и ниже по течению (76/86 нуклеотидов). [24]Alt1 разрушает стебель P2 в активной конформации, при этом предполагается, что P2 играет активирующую роль как для геномного, так и для антигеномного рибозима. [24] [27] [28] Alt2 представляет собой взаимодействие между вышестоящей фланкирующей последовательностью и рибозимом, а Alt3 представляет собой ненативное взаимодействие рибозим-рибозим. [24]
Вторичная структура этой ингибирующей конформации подтверждается различными экспериментальными подходами. [24] Сначала было проведено прямое зондирование с помощью рибонуклеаз, а последующее моделирование с помощью mfold 3.0 с использованием ограничений из результатов зондирования согласуется с предлагаемой структурой. [24] Во-вторых, ряд олигомеров ДНК, комплементарных различным областям AS1 / 2, был использован для восстановления активности рибозима; результаты подтверждают ингибирующую роль AS1 / 2. [24] В- третьих, мутационный анализ вводит одиночные / двойные мутации вне рибозима, чтобы убедиться, что наблюдаемая активность рибозима напрямую связана со стабильностью Alt1. [24] Было обнаружено, что стабильность AS1 обратно пропорциональна активности саморасщепления.[24]
Вторая пермиссивная структура позволяет рибозиму HDV саморасщепляться котранскрипционно, и эта структура дополнительно включает часть -54 / -18 нуклеотидов транскрипта РНК. [24] Находящийся выше ингибирующий участок -24 / -15 от вышеупомянутой ингибирующей конформации теперь изолирован в шпильке P (-1), расположенной выше сайта расщепления. [24] [29] [30] Мотив P (-1), однако, обнаруживается только в геномной последовательности, что может быть коррелировано с явлением, что геномные копии РНК HDV более многочисленны в инфицированных клетках печени. [24] [31]Экспериментальные данные также подтверждают эту альтернативную структуру. Во-первых, структурное картирование с помощью рибонуклеазы используется для зондирования фрагмента -54 / -1 вместо всего транскрипта-предшественника из-за быстро расщепляющейся природы этой структуры, которая согласуется с локальной шпилькой P (-1) (между -54 / -40 и -18 / -30 нт). [24] Во-вторых, эволюционная консервация обнаружена в P (-1) и связывающей области между P (-1) и P1 среди 21 геномного изолята РНК HDV. [24]
Использование в подготовке транскриптов РНК [ править ]
Особые свойства реакции расщепления рибозима HDV делают его полезным инструментом для получения РНК-транскриптов с гомогенными 3'-концами, альтернативой транскрипции РНК с помощью РНК-полимеразы Т7, которая часто может давать гетерогенные концы или нежелательные добавления. Вариант кДНК рибозима может быть получен рядом с кДНК последовательности РНК-мишени и РНК, полученной путем транскрипции с помощью РНК-полимеразы Т7. Последовательность рибозима будет эффективно расщепляться без дополнительных требований, поскольку -1 нуклеотид является инвариантным, оставляя 2'-3'-циклический фосфат, который можно легко удалить обработкой фосфатазой или полинуклеотидкиназой Т4. [32] Затем целевую РНК можно очистить с помощью гель-очистки.
Ссылки [ править ]
- ^ a b c d e f g h Ферре-Д'Амаре А. Р., Чжоу К., Дудна Д. А. (октябрь 1998 г.). «Кристаллическая структура рибозима вируса гепатита дельта». Природа . 395 (6702): 567–574. Bibcode : 1998Natur.395..567F . DOI : 10.1038 / 26912 . PMID 9783582 .
- ^ a b Modahl LE, Lai MM (июль 1998 г.). «Транскрипция мРНК антигена гепатита дельта продолжается на протяжении всей репликации вируса гепатита дельта (HDV): новая модель транскрипции и репликации РНК HDV» . Журнал вирусологии . 72 (7): 5449–5456. PMC 110180 . PMID 9621000 .
- ^ Макнотон TB, Ши ST, Modahl LE, Lai MM (апрель 2002). «Репликация РНК вируса гепатита дельта по катящемуся кругу осуществляется двумя различными клеточными РНК-полимеразами» . Журнал вирусологии . 76 (8): 3920–3927. DOI : 10,1128 / JVI.76.8.3920-3927.2002 . PMC 136092 . PMID 11907231 .
- ↑ Kuo MY, Sharmeen L, Dinter-Gottlieb G, Taylor J (декабрь 1988 г.). «Характеристика последовательностей саморасщепляющейся РНК на геноме и антигеноме дельта-вируса гепатита человека» . Журнал вирусологии . 62 (12): 4439–4444. PMC 253552 . PMID 3184270 .
- ↑ Chen PJ, Kalpana G, Goldberg J, Mason W, Werner B, Gerin J, Taylor J (ноябрь 1986). «Структура и репликация генома вируса гепатита дельта» . Труды Национальной академии наук Соединенных Штатов Америки . 83 (22): 8774–8778. DOI : 10.1073 / pnas.83.22.8774 . PMC 387014 . PMID 2430299 .
- ^ a b Webb CH, Lupták A (2011). «HDV-подобные саморасщепляющиеся рибозимы» . Биология РНК . 8 (5): 719–727. DOI : 10,4161 / rna.8.5.16226 . PMC 3256349 . PMID 21734469 .
- ^ Eickbush DG, Eickbush TH (июль 2010). «Ретротранспозоны R2 кодируют саморасщепляющийся рибозим для процессинга из котранскрипта рРНК» . Молекулярная и клеточная биология . 30 (13): 3142–3150. DOI : 10.1128 / MCB.00300-10 . PMC 2897577 . PMID 20421411 .
- ^ Webb CH, Riccitelli NJ, Ruminski DJ, Lupták A (ноябрь 2009). «Широкое распространение саморасщепляющихся рибозимов» . Наука . 326 (5955): 953. Bibcode : 2009Sci ... 326..953W . DOI : 10.1126 / science.1178084 . PMC 3159031 . PMID 19965505 .
- ^ Санчес-Лука FJ, Лопез MC, Масиас F, Алонсо C, Томас MC (октябрь 2011). «Идентификация рибозима, подобного вирусу гепатита дельта, на 5'-конце мРНК ретротранспозона L1Tc из Trypanosoma cruzi» . Исследования нуклеиновых кислот . 39 (18): 8065–8077. DOI : 10.1093 / NAR / gkr478 . PMC 3185411 . PMID 21724615 .
- ^ Санчес-Лука F, Лопез MC, Масиас F, Алонсо C, Томас MC (январь 2012). «Pr77 и L1TcRz: двойная система в 5'-конце ретротранспозона L1Tc, внутренний промотор и HDV-подобный рибозим» . Мобильные генетические элементы . 2 (1): 1–7. DOI : 10.4161 / mge.19233 . PMC 3383444 . PMID 22754746 .
- ^ Hetzel U, Szirovicza L, Smura T, Prähauser B, Vapalahti O, Kipar A, Hepojoki J (апрель 2019). «Идентификация нового дельтавируса у удавов» . mBio . 10 (2). DOI : 10,1128 / mBio.00014-19 . PMC 6445931 . PMID 30940697 .
- ^ Chang WS, Петерсон JH, Le Lay C, Ши M, Lo N, M Wille, Eden JS, Holmes EC (июль 2019). «Новые агенты, подобные гепатиту D, у позвоночных и беспозвоночных» . Эволюция вирусов . 5 (2): vez021. DOI : 10,1093 / ве / vez021 . PMC 6628682 . PMID 31321078 .
- ^ Перротта, AT; Бин, Мэриленд (14 января 1992 г.). «Расщепление олигорибонуклеотидов рибозимом, полученным из последовательности РНК вируса гепатита дельта». Биохимия . 31 (1): 16–21. DOI : 10.1021 / bi00116a004 . PMID 1731868 .
- ^ Ли TS, Радак BK, Harris ME, York DM (2016). «Путь конформационного переключения, опосредованного двумя металлами-ионами для активации рибозима HDV» . Катализ ACS . 6 (3): 1853–1869. DOI : 10.1021 / acscatal.5b02158 . PMC 5072530 . PMID 27774349 .
- ^ Gong B, Chen JH, Chase E, Chadalavada DM, Ядзима R, Golden BL, Bevilacqua PC, Кэри PR (октябрь 2007). «Прямое измерение pK (a), близкого к нейтральному, для каталитического цитозина в геномном рибозиме HDV с использованием рамановской кристаллографии». Журнал Американского химического общества . 129 (43): 13335–13342. DOI : 10.1021 / ja0743893 . PMID 17924627 .
- ^ а б Чен Дж. Х., Ядзима Р., Чадалавада Д. М., Чейз Е., Bevilacqua PC, Golden BL (август 2010 г.). «Кристаллическая структура 1,9 предварительного расщепления рибозима HDV предполагает, что как кислота Льюиса, так и общие кислотные механизмы вносят вклад в расщепление фосфодиэфира». Биохимия . 49 (31): 6508–6518. DOI : 10.1021 / bi100670p . PMID 20677830 .
- ^ a b Накано С., Проктор DJ, Bevilacqua PC (октябрь 2001 г.). «Механистическая характеристика геномного рибозима HDV: оценка каталитического и структурного вклада ионов двухвалентных металлов в механизм многоканальной реакции». Биохимия . 40 (40): 12022–12038. DOI : 10.1021 / bi011253n . PMID 11580278 .
- ^ Rajagopal P, Feigon J (июнь 1989). «Образование тройной цепи в гомопурине: гомопиримидиновые ДНК-олигонуклеотиды d (GA) 4 и d (TC) 4». Природа . 339 (6226): 637–640. DOI : 10.1038 / 339637a0 . PMID 2733796 .
- ^ Sklenár В, Feigon J (июнь 1990 г.). «Формирование стабильного триплекса из одиночной нити ДНК». Природа . 345 (6278): 836–838. Bibcode : 1990Natur.345..836S . DOI : 10.1038 / 345836a0 . PMID 2359461 .
- ^ Коннелл GJ, Ярус M (май 1994). «РНК с двойной специфичностью и двойные РНК со сходной специфичностью». Наука . 264 (5162): 1137–1141. Bibcode : 1994Sci ... 264.1137C . DOI : 10.1126 / science.7513905 . PMID 7513905 .
- ^ Лего P, Парди A (сентябрь 1994). «Исследование протонирования аденина в РНК на месте с помощью 13С ЯМР». Журнал Американского химического общества . 116 (18): 8390–8391. DOI : 10.1021 / ja00097a066 .
- ^ Каспрович А, Kempińska А, Smólska В, Wrzesiński Дж, Ciesiołka J (2015). «Применение флуоресцентно меченного транс-действующего антигеномного рибозима HDV для мониторинга взаимодействий антибиотик-РНК». Аналитические методы . 7 (24): 10414–10421. DOI : 10.1039 / C5AY02953H .
- ↑ Rosenstein SP, Been MD (октябрь 1991 г.). «Доказательства того, что элементы саморасщепляющейся геномной и антигеномной РНК вируса гепатита дельта имеют сходные вторичные структуры» . Исследования нуклеиновых кислот . 19 (19): 5409–5416. DOI : 10.1093 / NAR / 19.19.5409 . PMC 328906 . PMID 1923826 .
- ^ a b c d e f g h i j k l m n o p q Чадалавада Д. М., Кнудсен С. М., Накано С., Bevilacqua PC (август 2000 г.). «Роль вышестоящей структуры РНК в облегчении каталитической укладки рибозима геномного дельта-вируса гепатита». Журнал молекулярной биологии . 301 (2): 349–367. DOI : 10.1006 / jmbi.2000.3953 . PMID 10926514 .
- ^ Perrotta AT, Been MD (декабрь 1990). «Саморасщепляющийся домен геномной РНК вируса гепатита дельта: требования к последовательности и эффекты денатурирующего средства» . Исследования нуклеиновых кислот . 18 (23): 6821–6827. DOI : 10.1093 / NAR / 18.23.6821 . PMC 332737 . PMID 2263447 .
- ^ Perrotta AT, Been MD (апрель 1991). «Псевдоузелеподобная структура, необходимая для эффективного самоотщепления РНК вируса гепатита дельта». Природа . 350 (6317): 434–436. Bibcode : 1991Natur.350..434P . DOI : 10.1038 / 350434a0 . PMID 2011192 .
- ^ Matysiak M, Wrzesinski J, Ciesiołka J (август 1999). «Последовательная укладка геномного рибозима вируса гепатита дельта: структурный анализ промежуточных продуктов транскрипции РНК». Журнал молекулярной биологии . 291 (2): 283–294. DOI : 10.1006 / jmbi.1999.2955 . PMID 10438621 .
- ^ Perrotta AT, Никифорова O, Been MD (февраль 1999). «Консервированный выпуклый аденозин в периферическом дуплексе антигеномной саморасщепляющейся РНК HDV снижает кинетический захват неактивных конформаций» . Исследования нуклеиновых кислот . 27 (3): 795–802. DOI : 10.1093 / NAR / 27.3.795 . PMC 148249 . PMID 9889275 .
- ^ Mathews DH, Сабина J, Цукер M, Turner DH (Май 1999). «Расширенная зависимость термодинамических параметров от последовательности улучшает предсказание вторичной структуры РНК». Журнал молекулярной биологии . 288 (5): 911–940. DOI : 10.1006 / jmbi.1999.2700 . PMID 10329189 .
- ^ Цукер M, Mathews D, D Turner (1999). "Алгоритмы и термодинамика для предсказания вторичной структуры РНК". В Кларке JB (ред.). Биохимия и биотехнология РНК . Серия НАТО ASI. Дордрехт, Нидерланды: Kluwer Academic Publishers.
- ↑ Chen PJ, Kalpana G, Goldberg J, Mason W, Werner B, Gerin J, Taylor J (ноябрь 1986). «Структура и репликация генома вируса гепатита дельта» . Труды Национальной академии наук Соединенных Штатов Америки . 83 (22): 8774–8778. DOI : 10.1073 / pnas.83.22.8774 . PMC 387014 . PMID 2430299 .
- ^ Wichlacz A, Legiewicz M, Ciesiołka J (февраль 2004). «Создание транскриптов in vitro с гомогенными 3'-концами с использованием транс-действующего антигеномного дельта-рибозима» . Исследования нуклеиновых кислот . 32 (3): 39e – 39. DOI : 10.1093 / NAR / gnh037 . PMC 373431 . PMID 14973333 .
Внешние ссылки [ править ]
- Страница рибозима вируса гепатита дельта в Rfam
- Запись в базе данных субвирусной РНК для рибозима HDV