| KCNMA1 | |||||||
|---|---|---|---|---|---|---|---|
 Доменная структура каналов ВК | |||||||
| Идентификаторы | |||||||
| Символ | KCNMA1 | ||||||
| Альт. символы | SLO | ||||||
| Ген NCBI | 3778 | ||||||
| HGNC | 6284 | ||||||
| OMIM | 600150 | ||||||
| RefSeq | NM_002247 | ||||||
| UniProt | Q12791 | ||||||
| Прочие данные | |||||||
| Locus | Chr. 10 q22 | ||||||
| |||||||
{kind=link}
| KCNMB1 | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | KCNMB1 | ||||||
| Ген NCBI | 3779 | ||||||
| HGNC | 6285 | ||||||
| OMIM | 603951 | ||||||
| RefSeq | NM_004137 | ||||||
| UniProt | Q16558 | ||||||
| Прочие данные | |||||||
| Locus | Chr. 5 q34 | ||||||
| |||||||
| KCNMB2 | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | KCNMB2 | ||||||
| Ген NCBI | 10242 | ||||||
| HGNC | 6286 | ||||||
| OMIM | 605214 | ||||||
| RefSeq | NM_181361 | ||||||
| UniProt | Q9Y691 | ||||||
| Прочие данные | |||||||
| Locus | Chr. 3 кв. 26,32 | ||||||
| |||||||
| KCNMB3 | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | KCNMB3 | ||||||
| Альт. символы | KCNMB2, KCNMBL | ||||||
| Ген NCBI | 27094 | ||||||
| HGNC | 6287 | ||||||
| OMIM | 605222 | ||||||
| RefSeq | NM_171828 | ||||||
| UniProt | Q9NPA1 | ||||||
| Прочие данные | |||||||
| Locus | Chr. 3 кв. 26,3-кв. 27 | ||||||
| |||||||
| KCNMB3L | |
|---|---|
| Идентификаторы | |
| Символ | KCNMB3L |
| Альт. символы | KCNMB2L, KCNMBLP |
| Ген NCBI | 27093 |
| HGNC | 6288 |
| RefSeq | NG_002679 |
| Прочие данные | |
| Locus | Chr. 22 q11.1 |
| KCNMB4 | |||||||
|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||
| Символ | KCNMB4 | ||||||
| Ген NCBI | 27345 | ||||||
| HGNC | 6289 | ||||||
| OMIM | 605223 | ||||||
| RefSeq | NM_014505 | ||||||
| UniProt | Q86W47 | ||||||
| Прочие данные | |||||||
| Locus | Chr. 12 q15 | ||||||
| |||||||
| Альфа-субъединица калиевого канала ВК, активируемая кальцием | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | BK_channel_a | ||||||||
| Pfam | PF03493 | ||||||||
| ИнтерПро | IPR003929 | ||||||||
| |||||||||
Каналы BK (большой калий) представляют собой активируемые кальцием калиевые каналы с большой проводимостью , [1] также известные как Maxi-K , slo1 или Kca1.1 . Каналы BK - это калиевые каналы , управляемые напряжением, которые проводят большое количество ионов калия (K + ) через клеточную мембрану , отсюда и их название - большой калий . Эти каналы могут быть активированы (открыты) либо электрическими средствами, либо увеличением концентрации Ca 2+ в клетке. [2] [3] BK-каналы помогают регулировать физиологические процессы, такие какциркадные поведенческие ритмы и возбудимость нейронов. [4] BK-каналы также участвуют во многих процессах в организме, так как это распространенный канал. Они имеют тетрамерную структуру, которая состоит из трансмембранного домена , домена , чувствительного к напряжению, домена калиевого канала и цитоплазматического C-концевого домена , со многими рентгеновскими структурами для справки. Их функция - реполяризовать мембранный потенциал, позволяя калию вытекать наружу в ответ на деполяризацию или повышение уровня кальция.
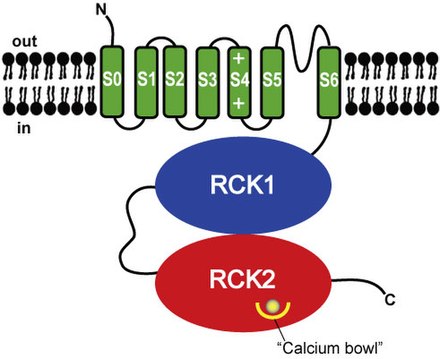
Структура [ править ]
Структурно BK-каналы гомологичны калиевым каналам , управляемым напряжением и лигандами , с датчиком напряжения и порами в качестве трансмембранного домена и цитозольного домена для связывания внутриклеточного кальция и магния . [5] Каждый мономер формирующей канал альфа-субъединицы является продуктом гена KCNMA1 (также известного как Slo1). Субъединица Slo1 имеет три основных структурных домена, каждый из которых выполняет свою функцию: домен измерения напряжения (VSD) определяет мембранный потенциал.через мембрану цитозольный домен (определяет концентрацию кальция, ионы Ca²) и домен поровых ворот (PGD), который открывается и закрывается для регулирования проникновения калия . Ворота активации находятся в PGD, который расположен либо на цитозольной стороне S6, либо на фильтре селективности (селективность - это предпочтение канала для проведения определенного иона). [5] Домен, чувствительный к напряжению, и домен, закрытый порами, вместе называются доменами, охватывающими мембрану, и образованы трансмембранными сегментами S1-S4 и S5-S6 соответственно. Внутри спирали S4 содержится ряд положительно заряженных остатков, которые служат датчиком первичного напряжения . [6]
Каналы BK очень похожи на каналы K⁺, управляемые напряжением, однако в каналах BK только один положительно заряженный остаток (Arg213) участвует в измерении напряжения через мембрану. [5] Также уникальным для каналов BK является дополнительный сегмент S0, этот сегмент требуется для модуляции β-субъединицы . [7] [8] и чувствительность к напряжению. [9]
Цитозольный домен состоит из двух доменов RCK (регулятор проводимости калия), RCK1 и RCK2. Эти домены содержат два сайта связывания Ca²⁺ с высоким сродством : один в домене RCK1, а другой в области, называемой чашей Ca², которая состоит из ряда остатков аспарагиновой кислоты (Asp), расположенных в домене RCK2. Сайт связывания Mg²⁺ расположен между VSD и цитозольным доменом, который образован: остатками Asp в петле S0-S1, остатками аспарагина в цитозольном конце S2 и остатками глутамина в RCK1. [5]При формировании сайта связывания Mg²⁺ два остатка происходят от RCK1 одной субъединицы Slo1, а два других остатка происходят от VSD соседней субъединицы. Для того, чтобы эти остатки координировали ион Mg²⁺, VSD и цитозольный домен из соседних субъединиц должны находиться в непосредственной близости. [5] Модуляторные бета-субъединицы (кодируемые KCNMB1 , KCNMB2 , KCNMB3 или KCNMB4 ) могут связываться с тетрамерным каналом . Существует четыре типа субъединиц β (β1-4), каждый из которых имеет разные паттерны экспрессии, которые изменяют стробирующие свойства BK-канала. Субъединица β1 в первую очередь отвечает за гладкомышечные клетки.При экспрессии субъединицы β2 и β3 экспрессируются нейронами, а β4 экспрессируются в головном мозге . [5] VSD ассоциируется с PGD посредством трех основных взаимодействий:
- Физическое соединение между VSD и PGD через линкер S4-S5.
- Взаимодействия между линкером S4-S5 и цитозольной стороной S6.
- Взаимодействия между S4 и S5 соседней субъединицы.
Регламент [ править ]
Каналы BK связаны и модулируются широким спектром внутри- и внеклеточных факторов, таких как вспомогательные субъединицы (β, γ), Slobs (slobs-связывающий белок), фосфорилирование , напряжение мембраны , химические лиганды (Ca²⁺, Mg²⁺), PKC. , BK α-субъединицы собираются 1: 1 с четырьмя различными вспомогательными типами β-субъединиц (β1, β2, β3 или β4). [10]
Было обнаружено, что перемещение и экспрессия BK каналов в плазматической мембране регулируется отдельными мотивами сплайсинга, расположенными внутри внутриклеточных C-концевых доменов RCK. В частности, вариант сплайсинга, в котором исключены эти мотивы, предотвращал экспрессию каналов ВК на клеточной поверхности и предполагает, что такой механизм влияет на физиологию и патофизиологию . [10]
BK-каналы в сосудистой системе модулируются веществами, которые естественным образом вырабатываются в организме, такими как ангиотензин II (Ang II), высокий уровень глюкозы или арахидоновая кислота (AA), которые модулируются при диабете за счет окислительного стресса (ROS). [10]
Более слабая чувствительность к напряжению позволяет каналам ВК работать в широком диапазоне мембранных потенциалов. Это гарантирует, что канал может правильно выполнять свою физиологическую функцию. [11]
Ингибирование активности BK-канала путем фосфорилирования S695 протеинкиназой C (PKC) зависит от фосфорилирования S1151 на С-конце альфа-субъединицы канала. Только одно из этих фосфорилирований в тетрамерной структуре должно произойти, чтобы ингибирование было успешным. Протеиновая фосфатаза 1 противодействует фосфорилированию S695. PKC снижает вероятность открытия канала за счет сокращения времени открытия канала и продления закрытого состояния канала. PKC не влияет на одноканальную проводимость, зависимость от напряжения или кальциевую чувствительность BK-каналов. [11]
Механизм активации [ править ]
Каналы ВК синергетически активируются за счет связывания ионов кальция и магния , но также могут активироваться через зависимость от напряжения. [10] Са²-зависимая активация происходит, когда внутриклеточный Ca²⁺ связывается с двумя сайтами связывания с высокой аффинностью : один расположен на С-конце домена RCK2 (чаша Ca²⁺), а другой - в домене RCK1. [5] Сайт связывания в домене RCK1 имеет несколько более низкое сродство к кальцию, чем чаша Ca², но отвечает за большую часть чувствительности к Ca²⁺. [12] Напряжение и кальций активируют каналы ВК, используя два параллельных механизма с датчиками напряжения.и сайты связывания Ca² соединяются с воротами активации независимо, за исключением слабого взаимодействия между двумя механизмами. Чаша из Ca² ускоряет кинетику активации при низких концентрациях Ca²⁺, в то время как сайт RCK1 влияет как на кинетику активации, так и дезактивацию. [11] Одна модель механизма была первоначально предложена Монодом, Вайманом и Ченжаксом, известная как модель MWC. Модель MWC для каналов BK объясняет, что конформационное изменение ворот активации при открытии канала сопровождается конформационным изменением сайта связывания Ca², что увеличивает аффинность связывания Ca². [12]
Магний-зависимая активация BK-каналов активируется через сайт связывания металла с низким сродством, который не зависит от Ca² Ca-зависимой активации. Датчик Mg²⁺ активирует каналы BK, сдвигая напряжение активации в более отрицательный диапазон. Mg²⁺ активирует канал только тогда, когда домен датчика напряжения остается в активированном состоянии. Домен цитозольного хвоста (CTD) представляет собой химический сенсор, который имеет несколько сайтов связывания для разных лигандов . CTD активирует канал BK при связывании с внутриклеточным Mg²⁺, чтобы обеспечить взаимодействие с доменом датчика напряжения (VSD). [11] Магний преимущественно координируется шестью атомами кислорода из боковых цепей кислородсодержащих остатков, карбонильных групп основной цепи.в белках или молекулах воды . [12] D99 на C-конце петли S0-S1 и N172 в петле S2-S3 содержат атомы кислорода боковой цепи в домене датчика напряжения, которые необходимы для связывания Mg²⁺. Подобно Ca²⁺-зависимой модели активации, Mg²-зависимая активация также может быть описана с помощью аллостерической модели стробирования MCW. В то время как кальций активирует канал в значительной степени независимо от датчика напряжения, магний активирует канал за каналом за счет электростатического взаимодействия с датчиком напряжения. [12] Это также известно как модель подталкивания, в которой магний активирует канал, толкая датчик напряжения посредством электростатических взаимодействий, и включает взаимодействия междубоковые цепи в различных структурных доменах. [5] Энергия, обеспечиваемая напряжением, связыванием Ca²⁺ и Mg²⁺, будет распространяться к активационным воротам каналов BK, чтобы инициировать ионную проводимость через поры. [5]
Воздействие на нейрон, орган, тело в целом [ править ]
Сотовый уровень [ править ]
Каналы BK помогают регулировать как возбуждение нейронов, так и высвобождение нейротрансмиттеров . [13] Эта модуляция синаптической передачи и электрического разряда на клеточном уровне происходит из-за экспрессии BK-канала в сочетании с другими калиево-кальциевыми каналами. [10] Открытие этих каналов вызывает привод в стороне равновесного потенциала калия и , следовательно , играет важную роль в ускорении реполяризации из потенциалов действия . [10] Это эффективно позволит более быструю стимуляцию. [10] Существует также роль в формировании общей реполяризации клеток, и, следовательно,после гиперполяризации (ГПД) потенциалов действия. [14] Роль, которую BK-каналы играют в быстрой фазе AHP, широко изучалась в гиппокампе. [14] Он также может играть роль в подавлении высвобождения нейротрансмиттеров. [15] Есть много BK каналов в клетках Пуркинье в мозжечке , тем самым подчеркивая их роль в координации движений и функции. [14] Кроме того, BK-каналы играют роль в модуляции активности дендритов, а также астроцитов и микроглии . [15] Они не только играют роль в ЦНС (центральная нервная система ), но также при сокращении гладких мышц , секреции эндокринных клеток и пролиферации клеток. [13] Различные субъединицы γ на раннем этапе развития мозга участвуют в возбудимости нейронов, а в невозбудимых клетках они часто являются движущей силой кальция. [10] Следовательно, эти субъединицы могут быть мишенями для терапевтического лечения в качестве активаторов BK-каналов. [10] Существуют дополнительные доказательства того, что ингибирование каналов BK может предотвратить отток калия и, таким образом, снизить использование АТФ , что, в сущности, обеспечивает выживание нейронов в условиях низкого содержания кислорода. [10]BK-каналы также могут действовать как нейрональный протектор, например, ограничивая проникновение кальция в клетки посредством окисления метионина . [10]
Уровень органа [ править ]
Каналы BK также играют роль в слушании . [14] Это было обнаружено, когда-субъединица BK была отключена у мышей и наблюдалась прогрессирующая потеря волосковых клеток улитки и, следовательно, потеря слуха. [14] BK-каналы участвуют не только в слухе, но и в циркадных ритмах . Slo-связывающие белки (Slobs) могут модулировать BK-каналы в зависимости от циркадных ритмов в нейронах. [10] BK-каналы экспрессируются в супрахиазматическом ядре (SCN), которое, как известно, влияет на патофизиологию сна. [14] Открыватели каналов BK также могут оказывать защитное действие насердечно-сосудистая система . [10] При низкой концентрации кальция каналы ВК в большей степени влияют на тонус сосудов . [10] Кроме того, сигнальная система каналов ВК в сердечно-сосудистой системе влияет на функционирование коронарного кровотока . [10] Одна из функций β-субъединицы в головном мозге включает ингибирование каналов ВК, что позволяет замедлить свойства каналов, а также помогает предотвратить судороги в височной доле . [10]
Уровень телесных функций [ править ]
Мутации BK-каналов, приводящие к снижению экспрессии мРНК , чаще встречаются у людей с умственными недостатками (из-за гипофункции [15] ), шизофреников или аутистов . [10] Более того, повышенная реполяризация, вызванная мутациями BK-каналов, может привести к алкогольной зависимости дискинезий , эпилепсии или пароксизмальных двигательных расстройств. [10] BK-каналы важны не только во многих клеточных процессах у взрослых, но и для правильного питания развивающегося плода . [10] Таким образом,эстроген может вызвать увеличение плотности каналов ВК в матке . [10] Однако в опухолевых клетках обнаружена повышенная экспрессия BK-каналов , и это может повлиять на будущую терапию рака , что более подробно обсуждается в разделе фармакологии. [10] ВК-каналы распространены по всему телу и, таким образом, оказывают большое и обширное влияние на организм в целом и на более клеточном уровне, как уже обсуждалось.
Фармакология [ править ]
Возможные проблемы [ править ]
Когда есть дефицит каналов BK, возникает ряд проблем. Последствия неисправности канала ВК могут повлиять на функционирование человека разными способами, некоторые из которых более опасны для жизни, чем другие. Каналы BK могут активироваться экзогенными загрязнителями и эндогенными газотрансмиттерами монооксидом углерода , [16] [17] оксидом азота и сероводородом. [18] Мутации в белках, участвующих в BK-каналах, или генах, кодирующих BK-каналы, участвуют во многих заболеваниях. Нарушение функции BK-каналов может распространяться при многих заболеваниях, таких как эпилепсия , рак , диабет , астма и гипертония.. [13] В частности, дефект β1 может повышать кровяное давление и задержку гидросалина в почках . [13] Мутации, связанные с утратой и усилением функции, вызывают такие расстройства, как эпилепсия и хроническая боль . [15] Кроме того, усиление активации BK-каналов за счет мутантов с избыточной функцией и амплификации связано с эпилепсией и раком. [13] Более того, BK-каналы играют роль как в опухолях, так и в раковых заболеваниях. При некоторых формах рака можно обнаружить gBK, вариантный ионный канал, называемый BK-каналом глиомы. [14] Известно, что каналы ВК каким-то образом влияют на деление клеток во время репликации., которые в нерегулируемом состоянии могут привести к раку и опухолям. [14] Более того, изученный аспект включает миграцию раковых клеток и роль, в которой каналы BK могут способствовать этой миграции, хотя многое еще неизвестно. [14] Другая причина важности понимания ВК-канала связана с его ролью в хирургии трансплантации органов . Это связано с активацией BK-каналов, влияющих на реполяризацию мембранного потенциала покоя . [10] Таким образом, понимание имеет решающее значение для безопасности эффективной трансплантации.
Текущие события [ править ]
Каналы ВК можно использовать в качестве фармакологических мишеней для лечения ряда заболеваний, включая инсульт [19] и гиперактивный мочевой пузырь . [20] Были попытки разработать синтетические молекулы, нацеленные на каналы BK, [21] однако их усилия пока оказались в значительной степени неэффективными. Например, BMS-204352, молекула, разработанная Bristol-Myers Squibb , не улучшила клинический исход у пациентов с инсультом по сравнению с плацебо . [22] Тем не менее, агонист BKCa каналов, BMS-204352, добился определенных успехов в лечении дефицита, наблюдаемого в Fmr1. нокаутные мыши , модель синдрома ломкой Х-хромосомы . [23] [24] BK-каналы также действуют как блокатор при ишемии и находятся в центре внимания при изучении их использования в качестве терапии инсульта. [10]
Будущие направления [ править ]
Есть много применений терапевтических стратегий с использованием BK-каналов. Было проведено исследование, показывающее, что блокада BK-каналов приводит к увеличению высвобождения нейротрансмиттеров, эффективно указывая на будущие терапевтические возможности в улучшении познания , улучшении памяти и облегчении депрессии . [13] Поведенческая реакция на алкоголь также модулируется BK-каналами, [10] поэтому дальнейшее понимание этой взаимосвязи может помочь в лечении пациентов, являющихся алкоголиками . Окислительный стресс на BK-каналах может привести к негативным последствиям снижения артериального давления за счет расслабления сердечно-сосудистой системы как на старение, так и на болезни.[10] Таким образом, сигнальная система может участвовать в лечении гипертонии и атеросклероза [10] посредством нацеливания на субъединицу для предотвращения этих пагубных эффектов. Кроме того, известная роль, которую BK-каналы могут играть при раке и опухолях, ограничена. Таким образом, в настоящее время не так много знаний о конкретных аспектах BK-каналов, которые могут влиять на опухоли и рак. [14] Дальнейшие исследования имеют решающее значение, поскольку они могут привести к огромному развитию методов лечения людей, страдающих от рака и опухолей. Известно, что эпилепсия возникает из-за чрезмерной возбудимости нейронов, BK-каналы которых имеют большое влияние на контроль сверхвозбудимости. [4]Следовательно, понимание может повлиять на лечение эпилепсии. В целом, каналы ВК являются мишенью для будущих фармакологических агентов, которые могут быть использованы для благоприятного лечения заболеваний.
См. Также [ править ]
- Субъединица альфа-1 калиевого канала, активируемого кальцием
- Калиевый канал, активируемый кальцием
- Калиевый канал с регулируемым напряжением
Ссылки [ править ]
- ↑ Zang K, Zhang Y, Hu J, Wang Y (2018). «Калийный канал с большой проводимостью и активированным напряжением (ВК) и эпилепсия». ЦНС и неврологические расстройства . 17 (4): 248–254. DOI : 10.2174 / 1871527317666180404104055 . PMID 29623857 .
- Перейти ↑ Miller, C. (2000). Геномная биология, 1 (4), обзоры 0004.1. https://dx.doi.org/10.1186/gb-2000-1-4-reviews0004
- Перейти ↑ Yuan, P., Leonetti, M., Pico, A., Hsiung, Y., & MacKinnon, R. (2010). Структура аппарата для активации Ca2 + -канала BK человека при разрешении 3,0 A. Наука, 329 (5988), 182-186. https://dx.doi.org/10.1126/science.1190414
- ^ a b N'Gouemo P (ноябрь 2011 г.). «Нацеливание на BK (большие калиевые) каналы при эпилепсии» . Мнение эксперта о терапевтических целях . 15 (11): 1283–95. DOI : 10.1517 / 14728222.2011.620607 . PMC 3219529 . PMID 21923633 .
- ^ a b c d e f g h i Ли США, Цуй Дж. (сентябрь 2010 г.). «Активация канала ВК: структурные и функциональные идеи» . Тенденции в неврологии . 33 (9): 415–23. DOI : 10.1016 / j.tins.2010.06.004 . PMC 2929326 . PMID 20663573 .
- ^ Аткинсон Н., Робертсон А., Ganetzky B (август 1991). «Компонент активированных кальцием калиевых каналов, кодируемых slo-локусом Drosophila». Наука . 253 (5019): 551–5. DOI : 10.1126 / science.1857984 . PMID 1857984 . S2CID 11317087 .
- ^ Morrow JP, Захаров С. И., Лю G, Ян L, Sok AJ, Маркс SO (март 2006). «Определение доменов канала BK, необходимых для модуляции бета1-субъединицы» . Труды Национальной академии наук Соединенных Штатов Америки . 103 (13): 5096–101. DOI : 10.1073 / pnas.0600907103 . PMC 1458800 . PMID 16549765 .
- ^ Валлнер M, P Меера, Toro L (декабрь 1996). «Детерминант регуляции бета-субъединицы в активируемых напряжением и Ca (2 +) -чувствительных K + каналах с высокой проводимостью: дополнительная трансмембранная область на N-конце» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (25): 14922–7. DOI : 10.1073 / pnas.93.25.14922 . PMC 26238 . PMID 8962157 .
- Перейти ↑ Koval OM, Fan Y, Rothberg BS (март 2007). «Роль трансмембранного сегмента S0 в зависимом от напряжения стробировании каналов ВК» . Журнал общей физиологии . 129 (3): 209–20. DOI : 10,1085 / jgp.200609662 . PMC 2151615 . PMID 17296928 .
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z Герман А., Ситдикова Г. Ф., Вейгер TM (август 2015 г.). «Окислительный стресс и макси-активированные кальцием калиевые (ВК) каналы» . Биомолекулы . 5 (3): 1870–911. DOI : 10.3390 / biom5031870 . PMC 4598779 . PMID 26287261 .
- ^ а б в г Ян Х, Чжан Г, Цуй Дж (2015). «Каналы ВК: несколько датчиков, один активатор» . Границы физиологии . 6 : 29. DOI : 10,3389 / fphys.2015.00029 . PMC 4319557 . PMID 25705194 .
- ^ a b c d Цуй Дж, Ян Х, Ли США (март 2009 г.). «Молекулярные механизмы активации ВК-каналов» . Клеточные и молекулярные науки о жизни . 66 (5): 852–75. DOI : 10.1007 / s00018-008-8609-х . PMC 2694844 . PMID 19099186 .
- ^ Б с д е е Ю. М., Лю SL, Sun PB, Pan H, Tian CL, Zhang LH (январь 2016). «Пептидные токсины и низкомолекулярные блокаторы ВК-каналов» . Acta Pharmacologica Sinica . 37 (1): 56–66. DOI : 10.1038 / aps.2015.139 . PMC 4722972 . PMID 26725735 .
- ^ a b c d e f g h i j Bentzen BH, Olesen SP, Rønn LC, Grunnet M (2014). «Активаторы каналов ВК и их терапевтические перспективы» . Границы физиологии . 5 : 389. DOI : 10,3389 / fphys.2014.00389 . PMC 4191079 . PMID 25346695 .
- ^ a b c d Contet C, Goulding SP, Kuljis DA, Barth AL (2016). «Каналы БК в центральной нервной системе». Big on Bk - Современное понимание функции K + -каналов с большой проводимостью, активированных напряжением и Ca2 +, на молекулярном, клеточном и системном уровнях . Международный обзор нейробиологии . 128 . С. 281–342. DOI : 10.1016 / bs.irn.2016.04.001 . ISBN 9780128036198. PMC 4902275 . PMID 27238267 .
- ^ Дюбюи Е, Потье М, Ван Р, Vandier С (февраль 2005 г.). «Непрерывное вдыхание окиси углерода ослабляет развитие гипоксической легочной гипертензии, предположительно за счет активации каналов BKCa» . Сердечно-сосудистые исследования . 65 (3): 751–61. DOI : 10.1016 / j.cardiores.2004.11.007 . PMID 15664403 .
- Перейти ↑ Hou S, Xu R, Heinemann SH, Hoshi T (март 2008 г.). «Высокоаффинный датчик Ca2 + RCK1 придает чувствительность к окиси углерода каналам Slo1 BK» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (10): 4039–43. DOI : 10.1073 / pnas.0800304105 . PMC 2268785 . PMID 18316727 .
- ^ Ситдиков GF, Weiger Т.М., Герман A (февраль 2010). «Сероводород увеличивает активность кальций-активированного калиевого (ВК) канала опухолевых клеток гипофиза крысы». Pflügers Archiv . 459 (3): 389–97. DOI : 10.1007 / s00424-009-0737-0 . PMID 19802723 . S2CID 23073556 .
- ^ Gribkoff В.К., Starrett JE, Dworetzky SI (апрель 2001). «Калиевые каналы Maxi-K: форма, функция и модуляция класса эндогенных регуляторов внутриклеточного кальция». Невролог . 7 (2): 166–77. DOI : 10.1177 / 107385840100700211 . PMID 11496927 . S2CID 8791803 .
- ^ Layne JJ, Nausch B, Олезен SP, Nelson MT (февраль 2010). «Активация канала ВК с помощью NS11021 снижает возбудимость и сократимость гладких мышц мочевого пузыря» . Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 298 (2): R378–84. DOI : 10,1152 / ajpregu.00458.2009 . PMC 2828174 . PMID 19923353 .
- ^ Gribkoff В.К., Winquist RJ (май 2005). "Модуляторы катионных каналов с регулируемым напряжением для лечения инсульта". Заключение эксперта по исследуемым препаратам . 14 (5): 579–92. DOI : 10.1517 / 13543784.14.5.579 . PMID 15926865 . S2CID 10236998 .
- Перейти ↑ Jensen BS (2002). «BMS-204352: открыватель калиевых каналов, разработанный для лечения инсульта» . Обзоры наркотиков ЦНС . 8 (4): 353–60. DOI : 10.1111 / j.1527-3458.2002.tb00233.x . PMC 6741660 . PMID 12481191 .
- ^ Laumonnier F, Roger S, Guérin P, Molinari F, M'rad R, Cahard D, Belhadj A, Halayem M, Persico AM, Elia M, Romano V, Holbert S, Andres C, Chaabouni H, Colleaux L, Constant J , Ле Геннек Дж. Ю., Брио С. (2006). «Связь функционального дефицита канала BKCa, синаптического регулятора возбудимости нейронов, с аутизмом и умственной отсталостью». Американский журнал психиатрии . 163 (9): 1622–1629. DOI : 10,1176 / ajp.2006.163.9.1622 . PMID 16946189 . S2CID 25225269 .
- ^ Hébert B; Pietropaolo S; Même S; Laudier B; Laugeray A; Doisne N; Квартал А; Lefeuvre S; Получил L; Cahard D; Laumonnier F; Crusio WE ; Pichon J; Менуэт А; Perche O; Брио С (2014). «Спасение фенотипов синдрома ломкой Х-хромосомы у мышей Fmr1 KO с помощью молекулы, открывающей канал BKCa» . Журнал "Орфанет редких болезней" . 9 : 124. DOI : 10,1186 / s13023-014-0124-6 . PMC 4237919 . PMID 25079250 .
Дальнейшее чтение [ править ]
- Ge L, Hoa NT, Wilson Z, Arismendi-Morillo G, Kong XT, Tajhya RB, Beeton C, Jadus MR (октябрь 2014 г.). «Большие ионные каналы калия (BK) в биологии, болезни и возможные цели для иммунотерапии рака» . Международная иммунофармакология . 22 (2): 427–43. DOI : 10.1016 / j.intimp.2014.06.040 . PMC 5472047 . PMID 25027630 .
- Кайл Б.Д., Браун А.П. (2014). «Регуляция активности ВК-каналов с помощью пре- и посттрансляционных модификаций» . Границы физиологии . 5 : 316. DOI : 10,3389 / fphys.2014.00316 . PMC 4141542 . PMID 25202279 .
- Нарди А, Олесен СП (2008). «Модуляторы каналов ВК: полный обзор». Современная лекарственная химия . 15 (11): 1126–46. DOI : 10.2174 / 092986708784221412 . PMID 18473808 .
- Чжан Дж, Ян Дж (2014). «Регулирование каналов ВК вспомогательными γ-субъединицами» . Границы физиологии . 5 : 401. DOI : 10,3389 / fphys.2014.00401 . PMC 4197896 . PMID 25360119 .
Внешние ссылки [ править ]
- Каналы BK + в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- «Активированные кальцием калиевые каналы» . База данных рецепторов и ионных каналов IUPHAR . Международный союз фундаментальной и клинической фармакологии.