{kind=link}

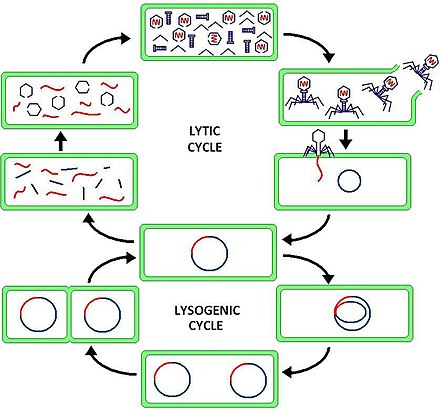
Лизогения , или лизогенный цикл , является одним из двух циклов вирусной репродукции ( литический цикл является другим). Лизогении характеризуются интеграцией бактериофага нуклеиновой кислоты в геном бактерии - хозяина или образование кругового репликона в бактериальной цитоплазме. В этом состоянии бактерия продолжает нормально жить и размножаться. Генетический материал бактериофага, называемый профагом , может передаваться дочерним клеткам при каждом последующем делении клеток, а при более поздних событиях (таких как УФ-излучение или присутствие определенных химических веществ) может высвобождать его, вызывая пролиферацию новых фагов через литический цикл. [1]Лизогенные циклы также могут происходить у эукариот , хотя метод включения ДНК полностью не изучен.
Разница между лизогенным и литическим циклами заключается в том, что в лизогенных циклах распространение вирусной ДНК происходит посредством обычного прокариотического размножения, тогда как литический цикл является более непосредственным, поскольку он приводит к очень быстрому созданию множества копий вируса и ячейка разрушена. Одно из ключевых различий между литическим циклом и лизогенным циклом состоит в том, что последний не лизирует клетку-хозяин сразу. [2] Фаги, которые реплицируются только через литический цикл, известны как вирулентные фаги, в то время как фаги, которые реплицируются с использованием как литического, так и лизогенного циклов, известны как умеренные фаги. [1]
В лизогенном цикле ДНК фага сначала интегрируется в бактериальную хромосому, чтобы произвести профаг. Когда бактерия размножается, профаг также копируется и присутствует в каждой из дочерних клеток. Дочерние клетки могут продолжать репликацию с присутствующим профагом, или профаг может покинуть бактериальную хромосому, чтобы инициировать литический цикл. [1] В лизогенном цикле ДНК хозяина не гидролизуется, но в литическом цикле ДНК хозяина гидролизуется в литической фазе.
Бактериофаги [ править ]
Бактериофаги - это вирусы, которые инфицируют и размножаются внутри бактерии. Умеренные фаги (такие как фаг лямбда ) могут воспроизводиться, используя как литический, так и лизогенный цикл. Через лизогенный цикл геном бактериофага не экспрессируется и вместо этого интегрируется в геном бактерии с образованием профага . [3] Поскольку генетическая информация бактериофага включена в генетическую информацию бактерий в качестве профага, бактериофаг пассивно реплицируется, когда бактерия делится с образованием дочерних бактериальных клеток. [3] В этом сценарии клетки дочерних бактерий содержат профаги и известны как лизогены.. Лизогены могут оставаться в лизогенном цикле в течение многих поколений, но могут переключаться на литический цикл в любое время посредством процесса, известного как индукция. [3] Во время индукции ДНК профага вырезается из бактериального генома, транскрибируется и транслируется для образования белков оболочки вируса и регулирования литического роста. [3]
Модельным организмом для изучения лизогении является фаг лямбда. Интеграция профага, поддержание лизогении, индукция и контроль вырезания генома фага при индукции подробно описаны в статье о фаге лямбда .
Компромисс фитнеса для бактерий [ править ]
Бактериофаги паразитируют, потому что они заражают своих хозяев, используют бактериальный механизм для размножения и, в конечном итоге, лизируют бактерии. Умеренные фаги могут приводить как к преимуществам, так и к недостаткам для своих хозяев через лизогенный цикл. Во время лизогенного цикла геном вируса включается в виде профага, а репрессор предотвращает репликацию вируса. Тем не менее, фаг умеренного климата может избежать репрессии, чтобы размножаться, производить вирусные частицы и лизировать бактерии. [4] Ускользание от репрессии фага в умеренном климате было бы недостатком для бактерий. С другой стороны, профаг может передавать геныкоторые повышают вирулентность хозяина и устойчивость к иммунной системе. Кроме того, продуцируемый профагом репрессор, который предотвращает экспрессию генов профага, придает иммунитет бактериям-хозяевам от литической инфекции родственными вирусами. [4]
Другая система, « арбитриум» , была недавно описана для бактериофагов, инфицирующих несколько видов Bacillus , в которой выбор между лизисом и лизогенией передается между бактериями посредством пептидного фактора. [5] [6]
Лизогенная конверсия [ править ]
При некоторых взаимодействиях между лизогенными фагами и бактериями может происходить лизогенная конверсия, которую также можно назвать фаговой конверсией. Это когда умеренный фаг вызывает изменение фенотипа инфицированных бактерий, что не является частью обычного фагового цикла. Изменения часто могут затрагивать внешнюю мембрану клетки, делая ее непроницаемой для других фагов или даже увеличивая патогенную способность бактерий для хозяина. Таким образом, умеренные бактериофаги также играют роль в распространении факторов вирулентности , таких как экзотоксины и экзоферменты, среди бактерий. Затем это изменение остается в геноме инфицированных бактерий, копируется и передается дочерним клеткам.
Выживание бактерий [ править ]
Было показано, что лизогенная конверсия способствует образованию биопленок у Bacillus anthracis [7]. Штаммы B. anthracis, излеченные от всех фагов, не могут образовывать биопленки, которые представляют собой прикрепленные к поверхности сообщества бактерий, которые позволяют бактериям лучше получать доступ к питательным веществам и выдерживать стрессы окружающей среды. [8] Помимо образования биопленок у B. anthracis , лизогенное преобразование Bacillus subtilis , Bacillus thuringiensis и Bacillus cereus продемонстрировало повышенную скорость или степень споруляции. [7] Споруляция производит эндоспоры., которые представляют собой метаболически спящие формы бактерий, которые обладают высокой устойчивостью к температуре, ионизирующему излучению, сушке, антибиотикам и дезинфицирующим средствам. [7]
Бактериальная вирулентность [ править ]
Также было показано, что невирулентные бактерии трансформируются в высоковирулентные патогены посредством лизогенного преобразования с факторами вирулентности, переносимыми лизогенным профагом. [9] Гены вирулентности, содержащиеся в профагах в виде отдельных автономных генетических элементов, известных как дебилы , дают бактериям преимущество, которое косвенно приносит пользу вирусу за счет увеличения выживаемости лизогена. [7]
Примеры:
- Corynebacterium diphtheriae продуцирует токсин дифтерии только тогда, когда она инфицирована фагом β. В этом случае ген, кодирующий токсин, переносится фагом, а не бактерией. [10]
- Vibrio cholerae - это нетоксичный штамм, который может стать токсичным, продуцируя токсин холеры , когда он инфицирован фагом CTXφ.
- Шигеллы дизентерия , которая производит дизентерию имеет токсины , которые попадают в две основных группы, Stx1 и stx2, чьи гены считаются частью генома ламбдовидных профагов .
- Streptococcus pyogenes , продуцируют пирогенный экзотоксин , полученный путем лизогенного преобразования, вызывающий лихорадку и скарлатину, скарлатину .
- Некоторые штаммы Clostridium botulinum , вызывающие ботулизм , экспрессируют ботулинический токсин из генов, передаваемых фагами.
Предотвращение лизогенной индукции [ править ]
Были предложены стратегии борьбы с некоторыми бактериальными инфекциями путем блокирования индукции профага (перехода от литического цикла к лизогенному) путем устранения агентов индукции in vivo . [9] Активные формы кислорода (АФК), такие как перекись водорода, являются сильными окислителями, которые могут разлагаться на свободные радикалы и вызывать повреждение ДНК бактерий, что приводит к индукции профага. [9] Одной из возможных стратегий борьбы с индукцией профага является использование глутатиона , сильного антиоксиданта, который может удалять свободные радикалы. [9]Другой подход может состоять в том, чтобы вызвать сверхэкспрессию репрессора CI, поскольку индукция профага происходит только при слишком низкой концентрации репрессора CI. [9]
Ссылки [ править ]
- ^ a b c Кэмпбелл и Рис (2005). Биология . Сан-Франциско: Пирсон. С. 338–339.
- ^ Лодиш; и другие. (2008). Молекулярная клеточная биология . Нью-Йорк: WH Freeman. С. 158–159.
- ^ а б в г Ватсон; и другие. (2008). Молекулярная биология гена . Колд-Спринг-Харбор, Нью-Йорк: Лаборатория Колд-Спринг-Харбор. С. 784–786.
- ^ а б Чен; и другие. (21 июня 2005 г.). «Популяционная приспособленность и регуляция генов Escherichia coli бактериальными вирусами» . PLOS Биология . 3 (7): e229. DOI : 10.1371 / journal.pbio.0030229 . PMC 1151598 . PMID 15984911 .
- Перейти ↑ Callaway, Ewen (2017). «Вы говорите о вирусе? Фаги отправляют химические сообщения» . Природа . DOI : 10.1038 / nature.2017.21313 . Архивировано 29 сентября 2019 года . Проверено 11 сентября 2019 .
- ^ Stokar-Avihail A, Tal N, Erez Z, Lopatina A, Sorek R. Широкое использование пептидной связи в фагах, заражающих почву и патогенные бактерии. Клетка-хозяин и микроб . 2019 8 мая; 25 (5): 746-55.
- ^ a b c d Луи-Шарль Фортье; и другие. (23 апреля 2013 г.). «Важность профагов для эволюции и вирулентности бактериальных патогенов» . Вирулентность . 4 (5): 354–65. DOI : 10.4161 / viru.24498 . PMC 3714127 . PMID 23611873 .
- ^ Наделл; и другие. (13 июля 2011 г.). «Фитнес-компромисс между местной конкуренцией и распространением биопленок холерного вибриона» . PNAS . 108 (34): 14181–14185. Bibcode : 2011PNAS..10814181N . DOI : 10.1073 / pnas.1111147108 . PMC 3161532 . PMID 21825170 .
- ^ a b c d e Кин, Эрик К. (14 декабря 2012 г.). «Парадигмы патогенеза: нацеливание на мобильные генетические элементы болезни» . Границы клеточной и инфекционной микробиологии . 2 : 161. DOI : 10.3389 / fcimb.2012.00161 . PMC 3522046 . PMID 23248780 .
- ↑ Мокроусов I (январь 2009 г.). «Corynebacterium diphtheriae: разнообразие генома, популяционная структура и перспективы генотипирования». Инфекция, генетика и эволюция . 9 (1): 1–15. DOI : 10.1016 / j.meegid.2008.09.011 . PMID 19007916 .