| Saccharomycotina | |
|---|---|
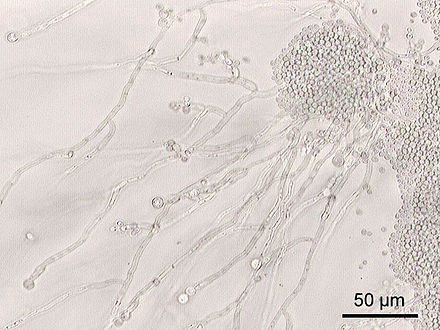 | |
| грибковые микроорганизмы албиканс | |
| Научная классификация | |
| Королевство: | Грибы |
| Разделение: | Аскомикота |
| Подразделение: | Сахаромикотина О.Е. Эрикс. И Winka 1997 [1] |
| Заказы / семьи | |
{kind=link}
Saccharomycotina - это подраздел (подтип) подразделения (типа) Ascomycota в Королевстве грибов . [2] [3] Он включает большинство дрожжей аскомицетов . Члены Saccharomycotina размножаются почкованием и не дают аскокарпов (плодовых тел). [2] [4]
Подразделение включает один класс: Saccharomycetes , который снова содержит один отряд: Saccharomycetales . [2] [3]
Заметными представителями Saccharomycotina являются пекарские дрожжи Saccharomyces cerevisiae и род Candida, который включает несколько патогенов человека.
Этимология [ править ]
Название происходит от греческого слова σάκχαρον ( sákkharon ), что означает «сахар», и μύκης ( mukēs ), что означает «гриб».
История и экономическое значение [ править ]
Исторические записи из Древнего Египта и Китая описывают процессы пивоварения и выпечки от 10 000 до 8 000 лет назад, а производство ферментированных напитков и продуктов, похоже, шло параллельно с возникновением сельского хозяйства . [5] В 1850-х годах Луи Пастер продемонстрировал, что дрожжи ответственны за ферментацию виноградного сока в вино. [6] [7]
Saccharomycotina включает некоторые из наиболее важных с экономической точки зрения известных грибов . Члены включают виды, имеющие промышленное и сельскохозяйственное значение (например, пивоварение , выпечка , ферментация пищевых продуктов, производство лимонной кислоты , производство рекомбинантных белков , производство биотоплива , биологическая борьба с вредителями сельскохозяйственных культур). Другие виды вызывают экономические потери во всем мире ( патогены растений , загрязнители продуктов питания и напитков). Третьи являются патогенами животных и человека . [8] [9]
Морфология [ править ]
Дрожжи-сахаромицеты обычно растут как одиночные клетки. Их клеточная морфология довольно проста, хотя форма их роста хорошо адаптирована. Аски голые, и аскоспоры могут иметь несколько форм. Аскокарпы (плодовые тела) не образуются ни у одного вида . Геномы сахаромицетов часто меньше геномов мицелиальных грибов . [2] [10] [11] [4]
Некоторые виды (например, виды Metschnikowia ) имеют тенденцию образовывать цепочки почкующихся клеток, которые называются псевдогифами . [2] Тем не менее, другие виды способны производить настоящие перегородчатые гифы . [4] Такие виды (например, Candida albicans ) называются диморфными, что означает, что они могут размножаться как почкующиеся дрожжи, так и как нитчатые гифы.
Воспроизведение [ править ]
Бесполое размножение [ править ]
Бесполое размножение происходит в основном вегетативно путем митоза и бутонизации . Saccharomycotina характеризуется холобластным почкованием [12], что означает, что все слои родительской клеточной стенки участвуют в событии почкования. Это оставляет шрам, через который больше не происходит почкование.
Бесполые клетки могут различаться по форме. [13] Форма клетки может быть информативной с точки зрения определения способа воспроизводства или таксономического отнесения к родам или видам.
Хотя это широко не известно, некоторые виды образуют эндоспоры (например, виды Candida ). [2] Это бесполые споры, которые образуются внутри их материнской клетки (гифы или отдельной клетки). Штаммы Candida и Metschnikowia также могут образовывать бесполые споры, называемые хламидоспорами . [2]
Половое размножение [ править ]
Половое размножение известно не для всех видов Saccharomycotina, но может происходить у некоторых видов, если условия окружающей среды благоприятствуют этому (например, дефицит азота и углеводов ). [2] Половое размножение хорошо известно у Saccharomyces cerevisiae . Здесь жизненный цикл включает в себя чередование гаплоидной и диплоидной фаз. Жизненный цикл протекает следующим образом: две клетки разного типа спаривания сливаются и ядра подвергаются кариогамии . Это приводит к дочерней клетке с диплоидным ядром, функционирующей как аск , где мейозпроисходит с образованием гаплоидных аскоспор . Когда аскоспоры прорастают, устанавливается гаплоидная фаза, которая поддерживается дальнейшим митозом и почкованием . В большинстве природных популяций эта фаза довольно короткая, поскольку аскоспоры сливаются почти сразу после возникновения мейоза . Это приводит к тому, что большинство популяций дрожжей диплоидны на протяжении большей части своего жизненного цикла. [4]
У Saccharomycotina есть два типа спаривания . Типы спаривания определяют пептидные гормоны, называемые феромонами, и соответствующие рецепторы для каждого типа. Эти феромоны организуют спаривание. Феромоны не влияют на один и тот же тип спаривания или диплоиды , но связываются с рецепторами разного типа спаривания. Взаимодействие между феромоном и рецептором приводит к изменению метаболизма, что позволяет осуществлять слияние между клетками разного типа спаривания. [4] [2]
Распространение и экология [ править ]
Сахаромицетные дрожжи встречаются почти во всех регионах мира, включая жаркие пустыни, полярные районы, пресную воду, соленую воду и атмосферу. [2] Их рост в основном сапротрофный , но некоторые из них являются важными патогенами растений и животных, включая человека. Они часто встречаются в специализированных средах обитания, например, в небольших объемах жидкости, богатой органическим углеродом (например, цветочного нектара). [4]
Примеры экологических режимов у Saccharomycotina:
- Ассоциации с насекомыми [14]
- Связь с растениями, включая Saccharomyces cerevisiae с виноградом [15]
- Паразитизм растений (например, гниль коробочек хлопка, вызванная Eremothecium ashbyi , Eremothecium gossypii как патоген на кофе, сое и других культурах) [2]
- Сапротрофизм на листьях и гниющей древесине (например, Ogataea ) [16]
- Патогены человека (например, виды Candida и Meyerozyma ) [8] [9]
Хотя дрожжи обычно изолированы от почвы, считается, что у немногих из них почва является основной средой обитания . [2]
Точная идентификация видов важна для понимания экологии дрожжей , что теперь стало возможным благодаря более широкому использованию методов на основе ДНК . До того, как стали доступны молекулярные методы, идентификация в основном основывалась на морфологии , что приводило к ошибочной классификации и в дальнейшем препятствовало получению надежных результатов экологических исследований.
Таксономия [ править ]
Saccharomycotina - это подраздел (подтип) подразделения (типа) Ascomycota . Это сестра группа для Pezizomycotina . [2] [3]
Дрожжи традиционно относили к отдельной группе грибного царства, но в последние годы [ когда? ] Методы на основе ДНК изменили понимание филогенетических взаимоотношений между грибами. Дрожжи считаются полифилетической группой [2] [11], состоящей из представителей Basidiomycota , Taphrinomycotina , а также Saccharomycotina. Это осознание привело к серьезным изменениям в филогении и таксономии Saccharomycotina. [2]
Кроме того, недавний [ когда? ] изменения в Международном кодексе номенклатуры водорослей, грибов и растений [17] [18] оказали большое влияние на классификацию грибов, включая Saccharomycotina. Изменения означают, что гриб может носить только одно правильное имя, т.е. отдельные имена для анаморфов и телеоморфов больше не допускаются. Это связано с серьезными изменениями в таксономии Saccharomycotina , поскольку многие виды в настоящее время описаны как на анаморфной, так и на телеоморфной стадиях. [18] Род Candida является примером рода, который подвергается масштабным пересмотрам.
Методы молекулярной идентификации являются важными инструментами для открытия новых видов и, следовательно, позволяют лучше понять биоразнообразие в этой группе. Большая часть будущей классификации Saccharomycotina будет основываться на филогенетическом анализе последовательностей ДНК , а не на морфологических характеристиках и признаках развития. [ необходима цитата ]
См. Также [ править ]
- Дрожжи в виноделии
- База данных генома сахаромицетов
Ссылки [ править ]
- Перейти ↑ Eriksson, OE & K. Winka (1997). «Надординальные таксоны Ascomycota ». Myconet . 1 : 1–16.
- ^ a b c d e f g h i j k l m n o Курцман, С. П., и Сугияма, Дж. (2015). «9 субклеточных структур и биохимических признаков в филогении грибов». Saccharomycotina и Taphrinomycotina: дрожжи и дрожжеподобные грибы Ascomycota. В Mycota: Комплексный трактат о грибах как экспериментальных системах для фундаментальных и прикладных исследований: VII Систематика и эволюция, часть B (2-е изд.). Берлин, Германия: Springer. С. 3–27. DOI : 10.1007 / 978-3-662-46011-5_9 . ISBN 978-3-662-46010-8.CS1 maint: несколько имен: список авторов ( ссылка )
- ↑ a b c Эрикссон, О. Э., и Винка, К. (1997). «Надординальные таксоны Ascomycota». Myconet .CS1 maint: несколько имен: список авторов ( ссылка )
- ^ Б с д е е Мур. Д., Робсон, Г. Д., Тринчи, APJ (2011). Путеводитель по грибам 21 века (2-е изд.). Кембридж, Великобритания: Издательство Кембриджского университета. С. 200–202.CS1 maint: несколько имен: список авторов ( ссылка )
- ^ Джон П. Арнольд (2005) [1911]. Происхождение и история пива и пивоварения: от доисторических времен до начала пивоваренной науки и технологий . Кливленд, Огайо: BeerBooks. п. 34. ISBN 978-0-9662084-1-2. OCLC 71834130 .
- ^ Пастер, Л. (1858). "Nouveaux faits about l'histoire de la fermentation alcoolique". Annales de Chimie et de Physique . 3 : 404–414.
- Перейти ↑ Manchester, KL (2007). «Луи Пастер, брожение и соперник». Южноафриканский научный журнал . 103 (9–10): 377–380.
- ^ a b Мартинс, Н., Феррейра, И.К., Баррос, Л., Сильва, С., Энрикес, М. (июнь 2014 г.). «Кандидоз: предрасполагающие факторы, профилактика, диагностика и альтернативное лечение». Микопатология . 177 (5–6): 223–240. DOI : 10.1007 / s11046-014-9749-1 . hdl : 1822/31482 . PMID 24789109 . S2CID 795450 . CS1 maint: несколько имен: список авторов ( ссылка )
- ↑ a b Эрдоган, А., Рао, СС (апрель 2015 г.). «Грибковое разрастание тонкой кишки». Curr Gastroenterol Rep . 17 (4): 16. DOI : 10.1007 / s11894-015-0436-2 . PMID 25786900 . S2CID 3098136 . CS1 maint: несколько имен: список авторов ( ссылка )
- ^ Мартин, Ф. (2014). Экологическая геномика грибов (1-е изд.). США: John Wiley & Sons, Inc.
- ^ a b Куртцман, К., Фелл, Дж. У., и Бекхаут, Т. (2011). Дрожжи: таксономическое исследование (5-е изд.). Амстердам, Нидерланды: Эльзевир.CS1 maint: несколько имен: список авторов ( ссылка )
- ^ фон Аркс, JA, и ван дер Вальт, JP (1987). Ophiostomatales и endomycetales. В: de Hoog GS, Smith MT, Weijman ACM (eds) Расширяющееся царство дрожжеподобных грибов . Амстердам, Нидерланды: Эльзевир.CS1 maint: несколько имен: список авторов ( ссылка )
- Перейти ↑ Kirk, PM, Cannon, PF, Minter, DW, Stalpers, JA (2008). Словарь грибов Эйнсворт и Бисби (10-е изд.). Уоллингфорд: CAB International.CS1 maint: несколько имен: список авторов ( ссылка )
- Перейти ↑ Vega, FE, and Blackwell, M. (2005). Насекомо-грибные ассоциации: экология и эволюция . Оксфорд, Великобритания: Издательство Оксфордского университета.CS1 maint: несколько имен: список авторов ( ссылка )
- ^ Мортимер, Р. и Polsinelli, М. (1999). «О происхождении винных дрожжей». Res Microbiol . 150 (3): 199–204. DOI : 10.1016 / s0923-2508 (99) 80036-9 . PMID 10229949 . CS1 maint: несколько имен: список авторов ( ссылка )
- ^ де Конинг, В., и Хардер, В. (1992). Метанол-утилизирующие дрожжи. В: Murell JC, Dalton H (eds) Утилизаторы метана и метанола . Нью-Йорк, США: Пленум. С. 207–244.CS1 maint: несколько имен: список авторов ( ссылка )
- ^ McNeill, J .; и другие. (2012). «Международный кодекс номенклатуры водорослей, грибов и растений». Regnum Vegetabile . 154 .
- ^ a b Хоксворт, DL (2011). «Новый рассвет для присвоения названий грибам: влияние решений, принятых в Мельбурне в июле 2011 г., на будущую публикацию и регулирование названий грибов» . IMA Fungus . 2 (2): 155–162. DOI : 10,5598 / imafungus.2011.02.02.06 . PMC 3359813 . PMID 22679600 .
Внешние ссылки [ править ]
- Древо жизни: Saccharomycetales