Миелиногенез - это, как правило, разрастание миелиновых оболочек в нервной системе и, в частности, прогрессирующая миелинизация нервных волокон аксонов в центральной нервной системе . Это неодновременный процесс, который происходит в основном постнатально у видов млекопитающих, начиная с эмбриона в разгар раннего развития и заканчивая после рождения. [1]

Функция [ править ]
Процесс миелинизации позволяет нейронным сигналам быстрее распространяться по аксону без потери сигнала. Это обеспечивает лучшую связь в определенных областях мозга, а также улучшает более широкие нейронные пути, соединяющие пространственно отдельные области, необходимые для многих сенсорных, когнитивных и моторных функций.
Некоторые ученые считают миелинизацию ключевым эволюционным преимуществом человека, обеспечивающим более высокую скорость обработки данных, что ведет к дальнейшей специализации мозга. Миелинизация продолжается еще как минимум 10–12 лет после рождения, прежде чем человек полностью разовьется. [2] Хотя скорость, с которой развиваются отдельные дети, различается, последовательность развития одинакова для всех детей (с диапазоном возрастов для выполнения конкретных задач развития).
Этапы [ править ]
Олигодендроциты отвечают за создание миелиновых оболочек в центральной нервной системе, в то время как шванновские клетки отвечают за периферическую нервную систему. Есть «две стадии маркеров OL: дифференциация OPCs в OL и окутывание аксонов…». [3]
Хотя механизмы и процессы миелинизации еще предстоит полностью понять, некоторые конкретные этапы этого процесса стали ясны:
- Этап 1: Аксонный контакт
- Этап 2: производство гена глиальных клеток
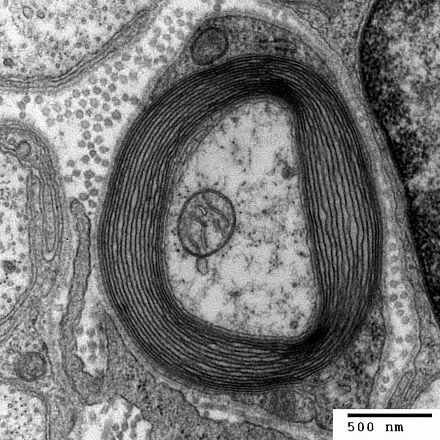
- Стадия 3: окантовка аксонов, которая является одной из двух фаз на ранних стадиях формирования миелиновой оболочки. Спиральное окутывание аксонов-мишеней начинается с выработки каждым инициирующим процессом ламеллярных расширений, которые проходят по окружности вокруг целевого аксона и тем самым формируют первый виток его миелиновой оболочки. [4]
- Этап 4: Созревание
.svg)
Механизм и процесс [ править ]
Основные молекулярные механизмы, которые контролируют процесс и последовательность миелиногенеза, полностью не известны. Многочисленные исследования были в первую очередь сосредоточены на упрощении основного нейронального контроля миелиногенеза, и такие исследования предоставили несколько возможностей.
Одно раннее исследование было сосредоточено на передаче сигналов о миелинизации олигодендроцитов путем регенерации периферических аксонов. Исследователи изучали регенерацию аксонов ПНС в течение 28 недель, чтобы выяснить, стимулируют ли периферические аксоны олигодендроциты для начала миелинизации. Экспериментальная индукция миелинизации путем регенерации периферических аксонов продемонстрировала, что шванновские клетки и олигодендроциты имеют общий механизм стимуляции миелинизации. [5] Аналогичное исследование, направленное на получение доказательств нейрональной регуляции миелиногенеза, показало, что образование миелина происходит из-за шванновских клеток, которые контролируются неопределенным свойством ассоциированного аксона. [5]
Другое такое исследование на мышах определило, что фактор транскрипции спираль-петля-спираль, OLIG1 , играет важную роль в процессе миелиногенеза олигодендроцитов. OLIG1 контролирует регуляцию некоторых генов, связанных с миелином, подавляя при этом другие. На клеточном уровне исследование экспериментально продемонстрировало, что OLIG1 необходим для стимуляции миелинизации олигодендроцитами в головном мозге. Однако олигодендроциты, относящиеся к спинному мозгу, продемонстрировали значительно меньшую потребность в регуляции OLIG1 для начала миелинизации. [6]
Недавние исследования на крысах показали, что раздельное действие апотрансферрина и гормона щитовидной железы может играть важную роль в миелинизации. Более того, исследования также показывают, что функция трансферрина и гормона щитовидной железы вместе контролирует миелиногенез.
В первую очередь, это исследование показало, что из-за гипертиреоза, который возник в результате увеличения экспрессии трансферрина, а также апотрансферрин-зависимой регуляции рецептора гормона щитовидной железы альфа , трансферрин, вероятно, был связан с эффектами гормона щитовидной железы на созревание олигодендроцитов и, в конечном итоге, миелинизацию.
Считается, что эта связь имеет место на уровне рецепторов гормонов щитовидной железы. Иммуногистохимический анализ был использован для дальнейшего подтверждения взаимосвязи между гормоном щитовидной железы и апотрансферрином во время олигодендрогенеза. [7]
{kind=link}
{kind=link}
Исследования по контролю миелиногенеза [ править ]
Исследования показали, что миелиногенез контролируется синтезом белков P1, P2 и P0. [8] Используя SDS-PAGE , исследователи выявили отдельные полосы с размерами полос 27 000 дальтон (P1), 19 000 дальтон (P2) и 14 000 дальтон (P0). Исследования также показали, что P1 и P2 активны до Po, поскольку этот белок поступает из периферической нервной системы. [8] В процессе регенерации шванновские клеткиповторно синтезируют белки, связанные с миелин-специфическими белками, когда присутствие аксонов восстанавливается. Ключевой частью эксперимента было показать, что шванновские клетки не синтезируют миелиновые белки в отсутствие аксонов. Синтез определяемых миелин-специфических белков не происходил в шванновских клетках, свободных от аксонов, что доказывает, что аксоны инструктируют шванновские клетки инициировать продукцию миелиновых белков сразу после установления аксональной ассоциации. [8]
Хотя аксон полностью контролировал синтез этих белков, одного аксона недостаточно для миелиногенеза. По мере продвижения эксперимента становилось все более очевидным, что межмембранные взаимодействия между аксонами каким-то образом способствовали синтезу основных белков P1, P2 и P0. Вероятно, что аксоны миелинизированных волокон способны индуцировать синтез миелинового белка шванновских клеток на протяжении всей взрослой жизни.
Миелиногенез в зрительном нерве [ править ]
Процесс и механистическая функция миелиногенеза традиционно изучались с использованием ультраструктуры и биохимических методов на зрительных нервах крыс . Реализация этого метода исследования давно позволила экспериментально наблюдать миелиногенез в нервном нерве модельного организма, который полностью состоит из немиелинизированных аксонов. Кроме того, использование зрительного нерва крысы помогло исследователям раннего миелиногенеза понять неправильное и атипичное течение миелиногенеза. [9]
Одно раннее исследование показало, что в развивающихся зрительных нервах крысы образование олигодендроцитов и последующая миелинизация происходят постнатально . В зрительном нерве клетки олигодендроцитов делились в последний раз через пять дней, при этом начало образования миелина происходило примерно на 6 или 7 день или около того. Однако точный процесс, с помощью которого олигодендроциты стимулировались для производства миелина, еще не был полностью завершен. понятно, но ранняя миелинизация зрительного нерва была связана с увеличением производства различных липидов - холестерина, цереброзида и сульфатида. [9]
Когда исследователи начали проводить послеродовые исследования, они обнаружили, что миелиногенез в зрительном нерве крысы первоначально начинается с аксонов самого большого диаметра, а затем распространяется на оставшиеся более мелкие аксоны. На второй неделе после рождения образование олигодендроцитов замедлилось - к этому моменту 15% аксонов были миелинизированы - однако миелиногенез продолжал быстро увеличиваться. В течение четвертой послеродовой недели почти 85% аксонов зрительного нерва крысы были миелинизированы. [9] В течение пятой недели и ближе к шестнадцатой неделе миелинизация замедлялась, и оставшиеся немиелинизированные аксоны были заключены в миелин. [10] Через зрительный нерв крысы ранние исследования внесли значительный вклад в знания в области миелиногенеза.
Важность сульфата [ править ]
Исследования развивающегося зрительного нерва показали, что галактоцереброзид (который образует сульфатид) появляется на 9-й постнатальный день и достигает пика на 15-й постнатальный день. [9] Это выражение было похоже на период, когда зрительный нерв показал максимальный период миелинизации аксона. Поскольку активность миелинизации аксонов снижалась, можно было сделать вывод, что активность фермента параллельна включению сульфата ([35S]) в сульфатид in vivo.
Исследования зрительного нерва крысы показали, что через 15 дней после рождения наблюдается усиление миелинизации. До этого периода времени большинство аксонов, примерно 70%, не миелинизированы. В это время [35S] сульфат был включен в сульфатид, и активность цереброзида , сульфотрансферазы достигла пика активности фермента. Этот временной интервал также показал период максимальной миелинизации на основании биохимических данных. [9]
В центральной нервной системе сульфатид , сульфатированные гликопротеины и сульфатированные мукополисахариды, по-видимому, связаны с нейронами, а не с миелином. При построении графика количества сульфатида, полученного из [35S], и активности сульфотрансферазы, мы получаем выделенные пики. [9] Пики приходятся на 15-й послеродовой день. Эти пики соответствовали максимальному периоду миелинизации зрительного нерва, который наблюдался на протяжении всего эксперимента. [9]
Таким образом, ранняя фаза миелинизации коррелировала с повышением синтеза липидов, холестерина, цереброзида и сульфатида. [9] Вероятно, что эти соединения синтезируются и упаковываются в Аппарат Гольджи олигодендроглии. [9] Несмотря на то, что транспорт этих липидов неизвестен, похоже, что миелинизация задерживается без их синтеза.
Клиническое значение [ править ]
Поскольку миелин образует электрически изолирующий слой, который окружает аксон некоторых нервных клеток , любое демиелинизирующее заболевание может повлиять на функционирование нервной системы . Одним из таких заболеваний является рассеянный склероз (РС), при котором демиелинизация происходит в центральной нервной системе (ЦНС). [11] Хотя проводятся исследования по защите олигодендроцитов и стимулированию ремиелинизации при рассеянном склерозе , [12] современные методы лечения в основном направлены на роль иммунной системы в демиелинизации. [13]
История исследований [ править ]
Другой исследователь, Пол Флехсиг, провел большую часть своей карьеры, изучая и публикуя детали процесса в коре головного мозга человека. Обычно это происходит между двумя месяцами до и после родов . Он выделил 45 отдельных областей коры и фактически нанес на карту кору головного мозга по паттерну миелинизации. Первая область коры головного мозга для миелинизации находится в моторной коре (часть области Бродмана 4), вторая - обонятельная кора, а третья - часть соматосенсорной коры (BA 3,1,2).
Последними миелинизирующими областями являются передняя поясная кора (F # 43), нижняя височная кора (F # 44) и дорсолатеральная префронтальная кора (F # 45).
В извилинах головного мозга, как и во всех других частях центральной нервной системы, нервные волокна развиваются не везде одновременно, а шаг за шагом в определенной последовательности, причем этот порядок событий особенно сохраняется в отношении появления мозгового слоя. субстанция. В извилинах головного мозга наложение мозгового вещества (миелинизация) в некоторых местах уже началось за три месяца до созревания плода, в то время как в других местах многочисленные волокна лишены мозгового вещества даже через три месяца после рождения. Порядок следования извилин регулируется законом, идентичным закону, который, как я показал, действует в отношении спинного мозга , продолговатого мозга., и мезоцефалон, и о чем можно сказать примерно так - что, говоря приблизительно, одинаково важные нервные волокна развиваются одновременно, а волокна разной важности развиваются одно за другим в последовательности, определяемой императивным законом (Основной закон миелогенеза). Формирование мозгового вещества в некоторых извилинах почти завершено, в то время как в некоторых оно даже не началось, а в других продвинулось лишь незначительно. [14]
Ссылки [ править ]
- ^ Eilam, R .; Бар-Лев, ДД; Левин-Зайдман, С .; Цури, М .; LoPresti, P .; Sela, M .; Arnon, R .; Ахарони, Р. (2014). «Олигодендрогенез и мейлиногенез во время постнатального развития влияние глатирамера ацетата». Глия . 62 (4): 649–665. DOI : 10.1002 / glia.22632 . PMID 24481644 .
- ^ Паус, Т., Zijdenbos, А., Ворсли, К., Коллинз, Д.Л., Блументэл J., Гидд, Ю.Н., ... & Эванс, AC (1999). Структурное созревание нервных путей у детей и подростков: исследование in vivo. Наука, 283 (5409), 1908-1911.
- ^ Уоткинс, Т., Mulinyawe, С., наждак, Б., Баррес, В. (2008). Четкие стадии миелинизации, регулируемые Y-секретазой и астроцитами, в системе сокультивирования быстро миелинизации ЦНС. 555-569
- Перейти ↑ Friedrich, VL., Hardy, RJ., (1996). Прогрессивное ремоделирование отростка олигодендроцитарного отростка во время миелиногенеза. 243-54.
- ^ a b Вайнберг, Э., и Спенсер, П. (1979). Исследования по контролю миелиногенеза. 3. Передача сигналов о миелинизации олигодендроцитов путем регенерации периферических аксонов. Brain Research, 162 (2), 273-279. DOI: 10.1016 / 0006-8993 (79) 90289-0
- ↑ Синь, М. (2005). Миелиногенез и распознавание аксонов олигодендроцитами в головном мозге не связаны у мышей Olig1-Null. Журнал неврологии, 25 (6), 1354-1365. DOI: 10.1523 / jneurosci.3034-04.2005
- ^ Marziali, Л. Н., Гарсиа, Cl, Pasquini, JM (2015). Трансферрин и гормон щитовидной железы вместе контролируют миелиногенез. Экспериментальная неврология. Vol 265. 129–141.
- ^ a b c Политис, MJ, Н. Стернбергер, Кэти Эдерл и Питер С. Спенсер. «Исследования по контролю миелиногенеза». Журнал неврологии 2.9 (1982): 1252-266.
- ^ a b c d e f g h i Теннекун, Дж., Коэн, СР., Прайс, DL., Маккхэнн, Дж. (1977). Миелиногенез зрительного нерва. Морфологический, авторадиографический и биохимический анализ. Журнал клеточной биологии, 72 (3), 604-616.
- ^ Dangata Ю., Кауфман, М. (1997). Миелиногенез в зрительном нерве гибридных мышей (C57BL x CBA) F1: морфометрический анализ, Европейский журнал морфологии, 35 (1), 3-18.
- ^ "Страница информации о рассеянном склерозе NINDS" . Национальный институт неврологических расстройств и инсульта . 19 ноября 2015. Архивировано из оригинала 13 февраля 2016 года . Проверено 6 марта +2016 . CS1 maint: обескураженный параметр ( ссылка )
- ^ Роджерс, Джейн М .; Робинсон, Эндрю П .; Миллер, Стивен Д. (2013). «Стратегии защиты олигодендроцитов и усиления ремиелинизации при рассеянном склерозе» . Открытие медицины . 86 (86): 53–63. PMC 3970909 . PMID 23911232 .
- ^ Cohen JA (июль 2009). «Новые методы лечения рецидивирующего рассеянного склероза» . Arch. Neurol . 66 (7): 821–8. DOI : 10,1001 / archneurol.2009.104 . PMID 19597083 .
- ^ Flechsig, Павел (1901-10-19). «Развитие (миелогенетическая) локализация коры головного мозга человека» . Ланцет . 158 (4077): 1028. DOI : 10.1016 / s0140-6736 (01) 01429-5 .