Членистоногая ногой является одной из форм сочлененного придатка из членистоногих , как правило , используется для прогулок . Многие из терминов, используемых для сегментов ног членистоногих (называемых подомерами ), имеют латинское происхождение, и их можно спутать с терминами, обозначающими кости: тазик ( тазобедренный сустав , множественное число тазиков ), вертел , бедро (множественное число бедер ), большеберцовая кость (множественное число большеберцовых костей ), tarsus (множественное число tarsi ), седалищная кость (множественное число ischia ),плюсна , запястье , дактиль (имеется в виду палец ), надколенник (множественное число надколенников ).
Гомологии сегментов ног между группами трудно доказать и являются источником многих аргументов. Некоторые авторы постулируют до одиннадцати сегментов на ногу для последнего общего предка ныне живущих членистоногих [1] , но современных членистоногих имеют восемь или меньше. Утверждается [2] [3], что предковая нога необязательно должна быть такой сложной и что другие события, такие как последовательная потеря функции Hox -гена , могут приводить к параллельному увеличению сегментов ноги.
У членистоногих каждый сегмент ноги сочленяется со следующим сегментом в шарнирном суставе и может сгибаться только в одной плоскости. Это означает, что требуется большее количество сегментов для достижения тех же видов движений, которые возможны у позвоночных животных, у которых есть шарнирно-шарнирные шарниры вращения у основания передних и задних конечностей. [4]
Бираминовый и однотонный [ править ]
{kind=link}
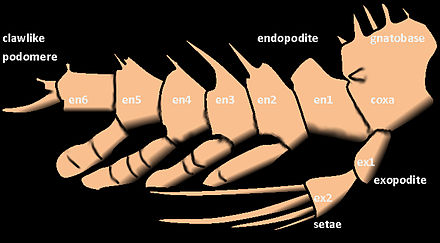
Придатки членистоногих могут быть как двуветвистыми, так и однообразными . Однородная конечность состоит из одной серии сегментов, прикрепленных встык. Однако двояковыпуклая конечность разветвляется на две части, и каждая ветвь состоит из ряда сегментов, прикрепленных встык.
Внешняя ветвь (ветвь) из придатков ракообразных известна как экзоподите или экзоподит , в то время как внутренняя ветвь известна как эндоподит или эндоподит . Другие структуры, помимо двух последних, называются exites (внешние структуры) и endites (внутренние структуры). Экзоподиты можно легко отличить от экзоподитов по наличию внутренней мускулатуры. Экзоподиты иногда могут отсутствовать у некоторых групп ракообразных ( амфиподы и равноногие ), а у насекомых они полностью отсутствуют. [5]
Ноги у насекомых и многоножек однообразны. У ракообразных первые усики однообразные, а вторые - двуветвистые, как и ноги у большинства видов.
Некоторое время считалось, что обладание однообразными конечностями является общим, производным признаком , поэтому однообразные членистоногие были сгруппированы в таксон под названием Uniramia . В настоящее время считается, что у нескольких групп членистоногих развились однообразные конечности независимо от предков с двуручными конечностями, поэтому этот таксон больше не используется.

Хелицерата [ править ]
Ноги паукообразных отличаются от таковых насекомых наличием двух сегментов с обеих сторон большеберцовой кости, надколенника между бедренной и большеберцовой костями и плюсневой кости (иногда называемой базитарсусом) между большеберцовой и предплюсневой костями (иногда называемой телотарзусом), в результате чего всего семь сегментов.
У лапок пауков есть когти на конце, а также крючок, который помогает при плетении паутины. Ноги паука также могут выполнять сенсорные функции, с волосками, которые служат рецепторами прикосновения, а также органом на предплюсне, который служит рецептором влажности, известным как орган предплюсны . [6]
Ситуация такая же у скорпионов , но с добавлением предплюсны за пределами предплюсны. Когти скорпиона на самом деле не ноги, а педипальпы , другой вид придатка, который также встречается у пауков и предназначен для хищничества и спаривания.
У Limulus нет плюсны или претарси, оставляя по шесть сегментов на каждой ноге.
Ракообразные [ править ]
Ноги ракообразных примитивно разделены на семь сегментов, которые не соответствуют системе именования, используемой в других группах. Это тазик, основание, седалищная кость, мерус, запястье, проподус и дактилюс. В некоторых группах некоторые сегменты конечностей могут срастаться. Коготь ( чела ) омара или краба образуется в результате сочленения дактиля с выростом проподуса. Конечности ракообразных также различаются двуветвью, тогда как у всех других современных членистоногих конечности однообразны.
Myriapoda [ править ]
Многоножки ( многоножки , многоножки и их родственники) имеют семь-сегментированные гуляющие ноги, включающие в себя тазик, вертел, prefemur, бедро, голень, лапку, и предплюсневую клешню. Ноги многоножек имеют множество модификаций в разных группах. У всех сороконожек первая пара ног видоизменена в пару ядовитых клыков, называемых форципулами. У большинства многоножек одна или две пары ходильных ног у взрослых самцов превращаются в структуры, передающие сперму, называемые гоноподами . У некоторых многоножек первая пара ног у самцов может быть уменьшена до крошечных крючков или окурков, в то время как у других первая пара может быть увеличена.
Насекомые [ редактировать ]
Насекомые и их родственники - это гексаподы, имеющие шесть ног, соединенных с грудной клеткой , каждая из которых состоит из пяти компонентов. По порядку от тела это тазик, вертел, бедро, большеберцовая кость и предплюсна. Каждый из них представляет собой один сегмент, за исключением лапки, которая может быть от трех до семи сегментов, каждый из которых называется тарсомером .
Фундаментальная морфология ног насекомых [ править ]
Типичная нога насекомого, например, у комнатной мухи или таракана , состоит из следующих частей, расположенных в последовательности от наиболее проксимальных к наиболее отдаленным :
- тазик
- вертел
- бедренная кость
- большеберцовая кость
- предплюсна
- претарсус.
С самой ногой связаны различные склериты вокруг ее основания. Их функции являются суставными и связаны с тем, как нога прикрепляется к основному экзоскелету насекомого. Такие склериты значительно различаются у неродственных насекомых. [7]
Coxa [ править ]
Тазик - это проксимальный сегмент и функциональная основа ноги. Он сочленяется с плевроном и связанными с ним склеритами грудного сегмента, а у некоторых видов он также сочленяется с краем стернита. Гомологии различных базальных склеритов открыты для обсуждения. Некоторые авторитеты предполагают, что они произошли от предкового подколенного сустава. У многих видов тазик имеет две доли, где он сочленяется с плеврой. Задняя доля - это мерон, который обычно составляет большую часть тазика. Мерон хорошо развит у Periplaneta, Isoptera, Neuroptera и Lepidoptera.
Trochanter [ править ]
Вертел сочленяется с тазиком, но обычно жестко прикреплен к бедренной кости. У некоторых насекомых его внешний вид может сбивать с толку; например, в Одонате есть два подсегмента. У паразитических перепончатокрылых основание бедра имеет вид второго вертела.
Бедренная кость [ править ]
У большинства насекомых бедро - самая большая часть ноги; это особенно заметно у многих насекомых с скачкообразными ногами, потому что типичный механизм прыжка заключается в выпрямлении сустава между бедренной и большеберцовой костью, а бедренная кость содержит необходимую массивную двупенистую мускулатуру.
Большеберцовая кость [ править ]
Большеберцовая кость - четвертая часть типичной ноги насекомого. Как правило, большеберцовая кость насекомого тонкая по сравнению с бедренной костью, но обычно она по крайней мере такая же, а часто и длиннее. Рядом с дистальным концом обычно имеется большеберцовая шпора, часто две или более. У Apocrita большеберцовая кость передней ноги несет большую апикальную шпору, которая перекрывает полукруглый промежуток в первом сегменте предплюсны. Щель покрыта гребешками, и насекомое очищает свои усики, протягивая их сквозь них.
Тарс [ править ]
Предковая лапка представляла собой единый сегмент, а у современных Protura , Diplura и некоторых личинок насекомых лапка также односегментирована. У большинства современных насекомых лапки разделены на части (тарсомеры), обычно около пяти. Фактическое количество зависит от таксона , что может быть полезно для диагностических целей. Например, Pterogeniidae обычно имеют 5-члениковые передние и средние лапки, но 4-члениковые задние лапки, тогда как у Cerylonidae есть четыре членика на каждой лапке.
Дистальный сегмент типичной ноги насекомого - это претарсус. У Collembola , Protura и многих личинок насекомых претарсус представляет собой одиночный коготь. На pretarsus большинство насекомых имеют пару когтей ( ungues , сингулярное ноготок ). Между ногтями расположена средняя пластина ангуитрактора, поддерживающая претарсус. Пластина прикрепляется к аподеме мышцы-сгибателя ногтевого сустава. У Neoptera паремподии представляют собой симметричную пару структур, отходящих от внешней (дистальной) поверхности пластинки ангуитрактора между когтями. [8] Он присутствует у многих Hemiptera и почти у всех Heteroptera . [8]Обычно паремподии щетинистые (щетинковидные), но у некоторых видов они мясистые. [9] Иногда паремподии уменьшаются в размерах и почти исчезают. [10] Над пластиной ангуитрактора претарсус расширяется вперед в среднюю долю, аролиум .
Webspinners ( Embioptera ) имеют увеличенный базальный членик на каждой из передних ног, содержащий железы , производящие шелк . [11]
Под претарсиями у представителей двукрылых обычно есть парные доли или пульвиллы, что означает «маленькие подушки». Под каждым ноготком находится по одному пульвиллу. Пульвиллы часто имеют аролиум между ними или срединную щетину или эмподиум , что означает место встречи пульвиллов. На нижней стороне сегментов предплюсны часто встречаются органы, похожие на пульвиллы или подошвы.. Arolium, plantulae и pulvilli - это адгезивные органы, позволяющие их обладателям подниматься по гладким или крутым поверхностям. Все они являются отростками экзоскелета, и их полости содержат кровь. Их структуры покрыты тонкими трубчатыми волосками, вершины которых увлажнены железистым секретом. Органы приспособлены для плотного прилегания волос к гладкой поверхности, так что адгезия происходит за счет поверхностных молекулярных сил. [7] [12]
Вариации функциональной анатомии ног насекомых [ править ]
Типичная грудная нога взрослого насекомого приспособлена для бега, а не для рытья, прыжков, плавания, хищничества или других подобных действий. Ноги большинства тараканов - хороший тому пример. Однако существует множество специализированных приспособлений, в том числе:
- Передние лапы Gryllotalpidae и некоторых Scarabaeidae приспособлены к рытью в земле.
- Передние лапы Mantispidae , Mantodea и Phymatinae приспособлены для захвата и удержания добычи одним способом, в то время как у Gyrinidae длинные ноги приспособлены для захвата пищи или добычи совсем другим способом.
- Передние лапы у некоторых бабочек, например у многих нимфалид , уменьшены настолько, что остаются только две пары функциональных ходильных ног.
- У большинства прямокрылых задние лапы скачкообразные ; у них сильно двупородные бедра и прямые длинные голени, приспособленные к прыжкам и, в некоторой степени, к защите ногами. Блохи, такие как представители подсемейства Halticinae, также имеют мощные задние бедра, которые позволяют им эффектно прыгать.
- Другие жуки с очень мускулистыми задними бедрами могут быть совсем не скачкообразными, но очень неуклюжими; например, определенные виды Bruchinae используют свои опухшие задние лапы, чтобы выбраться из семян растений с твердой оболочкой, таких как Erythrina, в которых они выросли до зрелого возраста.
- Ноги одоната , стрекоз и стрекоз приспособлены для захвата добычи, которой насекомые питаются во время полета или сидя на растении; они почти не могут использовать их для ходьбы. [7]
- Большинство водных насекомых используют свои ноги только для плавания, хотя многие виды незрелых насекомых плавают другими способами, такими как извивающиеся, волнообразные движения или выброс воды струями.
Эволюция и гомология ног членистоногих [ править ]
Эмбриональные сегменты тела ( сомиты ) различных таксонов членистоногих отклонились от простого строения тела с множеством сходных придатков, которые последовательно гомологичны, к разнообразию планов тела с меньшим количеством сегментов, оснащенных специализированными придатками. [13] Гомологии между ними были обнаружены путем сравнения генов в эволюционной биологии развития . [14]
| Сомит ( сегмент тела ) | Трилобит ( Trilobitomorpha ) | Паук ( Chelicerata ) | Сороконожка ( Myriapoda ) | Насекомое ( Hexapoda ) | Креветки ( ракообразные ) |
|---|---|---|---|---|---|
| 1 | усики | хелицеры (челюсти и клыки) | усики | усики | 1-й усик |
| 2 | 1-й этап | педипальпы | - | - | 2-й усик |
| 3 | 2-е ноги | 1-й этап | челюсти | челюсти | нижние челюсти (челюсти) |
| 4 | 3-е ноги | 2-е ноги | 1-я челюсть | 1-я челюсть | 1-я челюсть |
| 5 | 4 ноги | 3-е ноги | 2-я челюсть | 2-я челюсть | 2-я челюсть |
| 6 | 5 ноги | 4 ноги | воротник (без ног) | 1-й этап | 1-й этап |
| 7 | 6 ноги | - | 1-й этап | 2-е ноги | 2-е ноги |
| 8 | 7 ноги | - | 2-е ноги | 3-е ноги | 3-е ноги |
| 9 | 8 ноги | - | 3-е ноги | - | 4 ноги |
| 10 | 9 ноги | - | 4 ноги | - | 5 ноги |
Классификация модификаций ног [ править ]
- Ископаемые ноги приспособлены для рытья и рытья нор . Например: кротовины имеют окаменелые передние лапы.
- Saltatorial ноги модифицируются для прыжков и прыгаю . Например: у многих прямокрылых задние конечности скачкообразные.
- Нататорные ноги приспособлены для плавания . Например: у жуков- вихрей есть естественные среднегрудные и заднегруди.
- Бегущие ноги приспособлены для бега . Например: у многих тараканов очень чувствительные бегающие ноги.
- Хищные лапы приспособлены для хватания , обычно это охотничье приспособление. Например: богомолы используют свои хищные передние лапы для охоты.
Насекомые [ править ]
За исключением видов, у которых ноги были потеряны или стали рудиментарными в результате эволюционной адаптации, взрослые насекомые имеют шесть ног, по одной паре, прикрепленной к каждому из трех сегментов грудной клетки. У них есть парные придатки на некоторых других сегментах, в частности, на ротовом аппарате , усиках и церках , все из которых происходят от парных ног на каждом сегменте некоторого общего предка .
Однако у некоторых личинок насекомых есть дополнительные ходячие ноги на брюшных сегментах; эти дополнительные ноги называются ложноножками . Чаще всего они встречаются на личинках моли и пилильщиков. Ноги не имеют такого же строения, как ноги современных взрослых насекомых, и было много споров о том, гомологичны ли они им. [7] Текущие данные свидетельствуют о том, что они действительно гомологичны вплоть до очень примитивной стадии своего эмбриологического развития [15], но что их появление у современных насекомых не было гомологично между Lepidoptera и Symphyta . [16] Такие концепции широко распространены в современных интерпретациях филогении. [17]
В целом ноги личинок насекомых, особенно Endopterygota , различаются сильнее, чем у взрослых особей. Как уже упоминалось, у некоторых есть ложноножки, а также «настоящие» грудные ноги. У некоторых вообще нет видимых снаружи ног (хотя у них есть внутренние зачатки, которые при окончательном шелушении проявляются как взрослые ноги ). Примеры включают личинок мух или личинок долгоносиков . Напротив, у личинок других жесткокрылых , таких как Scarabaeidae и Dytiscidae, есть грудные ноги, но нет ложноножек. Некоторые насекомые, проявляющие гиперметаморфоз, начинают свой метаморфоз как планидии., специализированные активные личинки на ножках, но они заканчивают личиночную стадию как безногие личинки, например Acroceridae .
У Exopterygota ноги личинок имеют тенденцию напоминать ноги взрослых особей, за исключением приспособлений к их соответствующему образу жизни. Например, ноги большинства неполовозрелых Ephemeroptera приспособлены к тому, чтобы прятаться под подводными камнями и т.п., тогда как у взрослых особей ноги более изящные, и они не являются обузой во время полета. Опять же, детенышей Coccoidea называют «ползунками», и они ползают в поисках хорошего места для кормления, где они приживаются и остаются на всю жизнь. Их более поздние возрасты не имеют функциональных ног у большинства видов. У Apterygota ноги незрелых особей фактически являются уменьшенными версиями ног взрослых особей. [ необходима цитата]
Ссылки [ править ]
- ^ Kukalova-Пек, J. (1992). "Uniramia" не существует - план Pterygota, обнаруженный пермскими Diaphanopterodea из России (Insecta, Paleodictyopteroidea) ". Канадский зоологический журнал . 70 (2): 236–255. DOI : 10.1139 / z92-037 .
- ^ Фрайер, Г. (1996). «Размышления об эволюции членистоногих» . Биол. J. Linn. Soc . 58 (1): 1–55. DOI : 10.1111 / j.1095-8312.1996.tb01659.x .
- ^ Шрама, FR & S. Koenemann (2001). «Генетика развития и эволюция членистоногих: часть I, на ногах». Эволюция и развитие . 3 (5): 343–354. DOI : 10.1046 / j.1525-142X.2001.01038.x . PMID 11710766 .
- ^ Pat Willmer; Грэм Стоун; Ян Джонстон (12 марта 2009 г.). Экологическая физиология животных . Джон Вили и сыновья. п. 329. ISBN. 978-1-4443-0922-5.
- ^ Geoff А. Boxshall & Damia Жауме (2009). «Экзоподиты, эпиподы и жабры ракообразных» (PDF) . Систематика и филогения членистоногих . Museum für Tierkunde Dresden. 67 (2): 229–254. Архивировано из оригинального (PDF) 26 апреля 2019 года . Проверено 14 января 2012 .
- ^ Pechmann, Матиас (ноябрь 2010). «Механизмы формирования паттернов и морфологическое разнообразие придатков пауков и их значение для эволюции пауков» . Строение и развитие членистоногих . 39 (6): 453–467 . Проверено 20 августа 2020 .
- ^ a b c d Ричардс, О. У. Дэвис, Р.Г. (1977). Общий учебник энтомологии Иммса: Том 1: Структура, физиология и развитие Том 2: Классификация и биология . Берлин: Springer. ISBN 0-412-61390-5.
- ^ a b Фридеманн, Катрин; Спангенберг, Рико; Ёсизава, Казунор; Бойтель, Рольф Г. (2013). «Эволюция структур прикрепления в очень разнообразных Acercaria (Hexapoda)» (PDF) . Кладистика . 30 : 170–201. DOI : 10.1111 / cla.12030 . Архивировано из оригинального (PDF) 25 января 2014 года . Проверено 25 января 2014 года .
- ^ Шух, Randall Т. и Слейтер, Джеймс Александер (1995). Настоящие жуки мира (Hemiptera: Heteroptera): классификация и естествознание . Итака, Нью-Йорк: Издательство Корнельского университета. п. 46 . ISBN 978-0-8014-2066-5.
- ^ Гоел, SC (1972). «Заметки о строении пластинки ангуитрактора Heteroptera (Hemiptera)». Журнал энтомологии, Series A . 46 (2): 167–173. DOI : 10.1111 / j.1365-3032.1972.tb00124.x .
- ^ Росс, Эдвард С. (1991). «Эмбиоптера». В Наумане, ID; Карне, ПБ; и другие. (ред.). Насекомые Австралии. Том 1 (2-е изд.). Издательство Мельбурнского университета. С. 405–409.
- ↑ Станислав Н Горб. "Биологические приспособления для крепления: изучение разнообразия природы для биомиметиков Phil. Trans. R. Soc. A 2008; 366 (1870): 1557-1574 doi: 10.1098 / rsta.2007.2172 1471-2962
- ^ Фонд Новартис; Холл, Брайан (2008). Гомология . Джон Вили. п. 29. ISBN 978-0-470-51566-2.
- ^ Бруска, RC; Бруска, GJ (1990). Беспозвоночные . Sinauer Associates. п. 669 .
- ^ Панганибан, Грейс; Надя, Лиза; Кэрролл, Шон Б. «Роль гена Distal -less в развитии и эволюции конечностей насекомых». Текущая биология . 4 (8): 671–675. DOI : 10.1016 / S0960-9822 (00) 00151-2 .
- ^ Сузуки, Y; Палополи, MF (октябрь 2001 г.). «Эволюция брюшных придатков насекомых: являются ли ложноножки гомологичными или сходящимися?». Dev Genes Evol . 211 (10): 486–92. DOI : 10.1007 / s00427-001-0182-3 . PMID 11702198 .
- ^ Галис, Frietson (1996). «Эволюция насекомых и позвоночных: гены гомеобокса и гомология». Тенденции в экологии и эволюции . 11 (10): 402–403. DOI : 10.1016 / 0169-5347 (96) 30038-4 .