В эмбриологии стадия phylotypic или phylotypic период является определенной стадией развития или периода развития во середине эмбриогенеза , где зародыши родственных видов в пределах филюма выразить самую высокую степень морфологического и молекулярного сходства. Недавние молекулярные исследования на различных видах растений и животных позволили количественно оценить экспрессию генов, охватывающих важные стадии развития эмбриона, и обнаружили, что в течение морфологически определенного филотипического периода эволюционно самые старые гены, гены со сходными временными паттернами экспрессии и гены подвергаются сильнейшему очищающему отбору. наиболее активны на протяжении филотипического периода. [1]
Историческое происхождение концепции
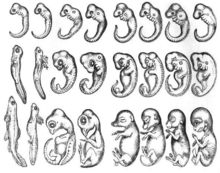

Идея о том, что эмбрионы разных видов имеют схожую морфологию на определенном этапе развития, восходит к Аристотелю . Аристотель наблюдал за рядом развивающихся эмбрионов позвоночных, отмечая в своем тексте «Поколение животных», что морфологические различия между разными эмбрионами возникли на поздних стадиях развития. В 1828 году Карл Эрнст фон Бэр создал свои законы эмбриологии , в которых обобщены результаты его сравнительных исследований эмбриогенеза . [2] В своем первом законе он предложил, чтобы более общие признаки группы появлялись в зародыше раньше, чем более специальные символы. [2] В 1866 году Эрнст Геккель.предположили, что каждый развивающийся организм проходит стадии эволюции своих предков, т. е. онтогенез повторяет филогенез . [3] Гипотеза о том, что разные организмы проходят стадии развития близкородственных организмов, устарела. Однако идея о том, что ранние стадии развития у видов сохраняются, с увеличением дивергенции по мере развития, повлияла на современную эволюционную биологию и биологию развития. [4] Ранняя консервационная или воронкообразная модель развития (см. Ниже) тесно связана с этим историческим происхождением.
Филотипический период
Первая формулировка концепции филотипического периода пришла в 1960 году из «Körpergrundgestalt» Фридриха Зайделя [5], что переводится как «основная форма тела». В 1977 году Коэн определил филетическую стадию как первую стадию, которая раскрывает общие черты, присущие всем членам этого типа. [6] Клаус Сандер пересмотрел эту концепцию в 1983 году и назвал ее филотипической стадией [7].что является «стадией наибольшего сходства между формами, которые в ходе эволюции по-разному специализировались как в своих образах взрослой жизни, так и в отношении самых ранних стадий онтогенеза». Обратите внимание, что это определение демонстрирует его поддержку модели песочных часов (см. В недавних работах филотипический период или филотипическая стадия рассматривается как период максимального сходства между видами в пределах каждого типа животных [8].
Хотя эта концепция была первоначально разработана с использованием морфологического сравнения развивающихся эмбрионов разных видов [7], период максимального сходства недавно был идентифицирован с использованием молекулярных данных. Филотипический период был идентифицирован с использованием сохранения экспрессии генов, [8] [9] оценок возраста гена, [10] [11] [12] сохранения последовательности гена, [13] экспрессии регуляторных генов и факторов транскрипции, [13] ] и взаимосвязь генов и белков. [14]
Модели воронок и песочных часов
Модель воронки - это гипотеза о том, что наиболее консервативная стадия развития (филотипический период) происходит в начале эмбриогенеза , с возрастающей дивергенцией по мере развития. Это также известно как ранняя консервационная модель развития.
Доказательства альтернативной модели возникли в результате тщательного сравнения временной дивергенции в морфологии эмбрионов разных видов. Например, Клаус Сандер заметил, что «невероятное разнообразие личинок и взрослых особей» насекомых происходит после того, как они «развиваются из почти идентичных зачатков на стадии зародышевого пояса». [7] Наиболее консервативная стадия развития, стадия зародышевой ленты, происходит ближе к середине развития, чем в начале, поддерживая средний период развития максимального сходства между видами. Эта модель, называемая моделью песочных часов [15] [16] , основана на идее о том, что ранние эмбрионы разных видов демонстрируют дивергентные формы, но их морфология сходится в середине развития, за которым следует период возрастающей дивергенции.
Поддержка модели песочных часов
В противоположность ранним морфологическим работам фон Бэра и Геккеля , недавние морфологические исследования продемонстрировали наибольшее расхождение среди близкородственных видов как на ранних стадиях развития ( гаструляция ), так и на поздних стадиях развития, [17] подтверждая модель песочных часов. Дальнейшая поддержка модели песочных часов пришла из открытия, что Hox-гены , группа последовательно активируемых генов, которые регулируют формирование передне-задней оси тела, активируются в середине развития на филотипической стадии. [18]Поскольку эти гены являются высококонсервативными и участвуют в формировании осей тела, активация Hox-генов может быть важным игроком в повышенной сохранности эмбрионов близкородственных видов во время среднего развития. [15]
Появление секвенирования следующего поколения позволило ученым использовать молекулярные методы для определения периода развития, в котором паттерны экспрессии генов наиболее консервативны среди различных видов. В 2010 году два исследования обнаружили молекулярные доказательства, подтверждающие модель песочных часов. [8] [10] Калинка и др . [8] секвенировали транскриптом шести видов Drosophila в течение периода развития, идентифицируя наиболее консервативную экспрессию генов в середине развития во время стадии развития зародышевой полосы членистоногих. Гены, которыми были обогащены развивающиеся эмбрионы на стадии зародышевого пояса, участвуют в клеточном и организменном развитии. Домазет-Лошо и Тауц [10]проанализировали транскриптом рыбок данио ( Danio rerio ) во время развития, от неоплодотворенных яиц до взрослых особей. Они использовали метод, называемый геномной филостратиграфией, чтобы оценить возраст каждого гена во время развития. У рыбок данио, а также в дополнительных наборах транскриптомных данных Drosophila , москита Anopheles и нематоды Caenorhabditis elegans авторы обнаружили, что гены, экспрессируемые во время среднего развития, старше, чем гены, экспрессируемые в начале и в конце развития, что подтверждает модель песочных часов.
Другие недавние геномные исследования подтвердили наличие филотипической стадии среднего развития у позвоночных [9] и у растения Arabidopsis thaliana . [11] [12] [19] Профили экспрессии временных генов для развивающейся мыши ( Mus musculus ), курицы ( Gallus gallus ), лягушки ( Xenopus laevis ) и рыбок данио ( Danio rerio ) показали, что наиболее консервативная экспрессия генов происходит у позвоночных. в середине развития на стадии глоточного зародыша. pharyngula Стадия наступает, когда у позвоночных развиваются четыре отличительных черты (хорда, спинной полый нервный шнур, постанальный хвост и серия парных жаберных щелей).
Поддержка модели раннего сохранения (воронки)
Недавние молекулярные данные также подтверждают раннюю модель сохранения. Например, Piasecka et al . [13] повторно проанализировали набор данных о рыбках данио, опубликованный Domazet-Lošo и Tautz. [10] Они обнаружили, что применение логарифмического преобразования к данным экспрессии генов изменило результаты, чтобы поддержать максимальную сохранность в раннем развитии. Кроме того, после объединения данных об экспрессии генов рыбок данио в «модули транскрипции», отражающие каждую стадию развития, они обнаружили несколько линий доказательств, подтверждающих раннюю модель сохранения (последовательность гена, возраст, размер семейства генов и сохранение экспрессии), в то время как только анализ регуляторные области генов поддерживали модель песочных часов. [13]
Одна из гипотез эволюционной консервации во время филотипического периода состоит в том, что это период, характеризующийся высоким уровнем взаимодействий, поскольку строится план тела. [14] У рыбок данио взаимосвязь белков в течение времени развития оказалась самой высокой на раннем этапе развития, что подтверждает раннюю модель сохранения. [14] Другой способ исследовать момент в развитии, в котором ограничения развития наиболее сильны, - это экспериментальная потеря гена, потому что удаление гена должно быть более вредным, когда он экспрессируется на стадии развития с более сильными эволюционными ограничениями. [20] Генный нокаутэксперименты на мышах и рыбках данио продемонстрировали, что соотношение основных генов к несущественным генам уменьшается с течением времени развития, предполагая, что существуют более сильные ограничения в раннем развитии, которые со временем ослабляются. [20] Несмотря на растущее количество доказательств, подтверждающих модель песочных часов, определение точки в развитии, которая наиболее сохраняется среди видов с типом (филотипический период), является противоречивым в области биологии развития.
Внутрифиловый и межфиловый филотипический период
Филотипический период определяется как период максимального сходства между видами с типом, но недавнее исследование сравнило филотипический период между разными типами, чтобы проверить, сохраняются ли одни и те же консервативные периоды во время развития в более глубоких филогенетических отношениях. Левин и др . [21] сравнили паттерны экспрессии генов в процессе развития у десяти особей из десяти разных типов животных и нашли доказательства модели обратных песочных часов дивергенции экспрессии генов между разными типами. [21] Эта модель обратных песочных часов отражает наблюдение, что экспрессия генов была значительно более дивергентной среди видов на переходном этапе среднего развития, в то время как экспрессия генов была более консервативной на ранних и поздних стадиях развития.[21] Хотя этот интригующий паттерн может иметь значение для нашего определения типа, [21] в последующем документе утверждается, что есть несколько методических проблем, которые необходимо решить, чтобы проверить гипотезу о том, что сроки ограничения развития различаются между филю по сравнению с филумом. Во-первых, сравнение одного представителя десяти различных типов могло бы отразить различия между типами, а также более глубокие или мелкие филогенетические ветви, которые попадают между этими десятью особями, поэтому необходима большая выборка внутри каждого типа. [22] Во-вторых, при парном сравнении каждый из десяти видов рассматривается как независимое наблюдение, но некоторые виды более близки, чем другие. [22]
Ссылки
- ^ Дрост, Хайк-Георг; Яница, Филипп; Гроссе, Иво; Квинт, Марсель (2017). «Сравнение разных королевств песочных часов развития» . Текущее мнение в области генетики и развития . 45 : 69–75. DOI : 10.1016 / j.gde.2017.03.003 . PMID 28347942 .
- ^ a b фон Бэр, Карл Эрнст (1828). Über Entwickelungsgeschichte der Thiere. Beobachtung und Reflexion . Кенигсберг: Bornträger.
- ^ Геккель, Эрнст (1866). Generelle Morphologie der Organismen . Берлин: Георг Реймер.
- ^ Сандер, Клаус; Шмидт-Отт, Урс (2004). «Эво-дево аспекты классических и молекулярных данных в исторической перспективе». Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития . 302B (1): 69–91. DOI : 10.1002 / jez.b.20003 . PMID 14760654 .
- Перейти ↑ Seidel, F. (1960). "Körpergrundgestalt und Keimstruktur. Eine Erörterung über die Grundlagen der vergleichenden und Experimentellen Embryologie und deren Gültigkeit bei phylogenetischen Berlegungen". Zoologischer Anzeiger . 164 : 245–305.
- ^ Коэн, Дж. (1977). Репродукция . Лондон: Баттерворт.
- ^ a b c Сандер, К. (1983). В развитии и эволюции: шестой симпозиум Британского общества биологии развития . Издательство Кембриджского университета.
- ^ a b c d Калинка, Алексей Т .; Варга, Каролина М .; Джеррард, Дэйв Т .; Прейбиш, Стефан; Коркоран, Дэвид Л .; Джарреллс, Джулия; Олер, Уве; Бергман, Кейси М .; Томанчак, Павел (09.12.2010). «Дивергенция экспрессии генов повторяет модель песочных часов развития». Природа . 468 (7325): 811–814. Bibcode : 2010Natur.468..811K . DOI : 10,1038 / природа09634 . ISSN 0028-0836 . PMID 21150996 . S2CID 4416340 .
- ^ а б Ириэ, Наоки; Куратани, Сигеру (22 марта 2011 г.). «Сравнительный анализ транскриптома выявляет филотипический период позвоночных в органогенезе» . Nature Communications . 2 : 248. Bibcode : 2011NatCo ... 2..248I . DOI : 10.1038 / ncomms1248 . ISSN 2041-1723 . PMC 3109953 . PMID 21427719 .
- ^ a b c d Домазет-Лошо, Томислав; Тауц, Дитард (09.12.2010). «Филогенетически основанный на транскриптоме возрастной индекс отражает паттерны онтогенетической дивергенции». Природа . 468 (7325): 815–818. Bibcode : 2010Natur.468..815D . DOI : 10,1038 / природа09632 . ISSN 0028-0836 . PMID 21150997 . S2CID 1417664 .
- ^ a b Квинт, Марсель; Дрост, Хайк-Георг; Габель, Александр; Ульрих, Кристиан Карстен; Бённ, Маркус; Гроссе, Иво (2012-10-04). «Транскриптомные песочные часы в эмбриогенезе растений». Природа . 490 (7418): 98–101. Bibcode : 2012Natur.490 ... 98Q . DOI : 10.1038 / nature11394 . ISSN 0028-0836 . PMID 22951968 . S2CID 4404460 .
- ^ a b Дрост, Хайк-Георг; Габель, Александр; Гроссе, Иво; Квинт, Марсель (2015-05-01). «Доказательства активного сохранения филотранскриптомических паттернов песочных часов в эмбриогенезе животных и растений» . Молекулярная биология и эволюция . 32 (5): 1221–1231. DOI : 10.1093 / molbev / msv012 . ISSN 0737-4038 . PMC 4408408 . PMID 25631928 .
- ^ a b c d Пясецка, Барбара; Лихоцкий, Павел; Моретти, Себастьян; Бергманн, Свен; Робинсон-Рехави, Марк (25 апреля 2013 г.). «Песочные часы и ранние модели сохранения - сосуществующие модели ограничений развития у позвоночных» . PLOS Genetics . 9 (4): e1003476. DOI : 10.1371 / journal.pgen.1003476 . ISSN 1553-7404 . PMC 3636041 . PMID 23637639 .
- ^ a b c Конт, Орели; Ру, Жюльен; Робинсон-Рехави, Марк (01.03.2010). «Молекулярная передача сигналов в развитии рыбок данио и филотипический период позвоночных» . Эволюция и развитие . 12 (2): 144–156. DOI : 10.1111 / j.1525-142X.2010.00400.x . ISSN 1525-142X . PMC 2855863 . PMID 20433455 .
- ^ a b Duboule, D. (1994-01-01). «Временная колинеарность и филотипическая прогрессия: основа стабильности позвоночного Бауплана и эволюции морфологии посредством гетерохронии». Разработка. Приложение : 135–142. PMID 7579514 .
- ^ Рафф, Рудольф А. (1996). Форма жизни: гены, развитие и эволюция формы животных . Издательство Чикагского университета. ISBN 9780226256573.
- ^ Шмидт, Кай; Старк, Дж. Матиас (2004-09-15). «Изменчивость в развитии во время раннего эмбрионального развития рыбок данио, Danio rerio». Журнал экспериментальной зоологии, часть B: Молекулярная эволюция и эволюция развития . 302B (5): 446–457. DOI : 10.1002 / jez.b.21010 . ISSN 1552-5015 . PMID 15580642 .
- ^ Slack, JMW; Голландия, PWH; Грэм, CF (1993-02-11). «Зотип и филотипический этап». Природа . 361 (6412): 490–492. Bibcode : 1993Natur.361..490S . DOI : 10.1038 / 361490a0 . PMID 8094230 . S2CID 4362531 .
- ^ Дрост, Хайк-Георг; Белльштедт, Джулия; Ó'Maoiléidigh, Diarmuid S .; Сильва, Андерсон Т .; Габель, Александр; Вайнхольдт, Клаус; Райан, Патрик Т .; Деккерс, Бас JW; Бенцинк, Леони; Хильхорст, Хенк WM; Ligterink, Wilco; Веллмер, Фрэнк; Гроссе, Иво; Квинт, Марсель (23 февраля 2016 г.). «Постэмбриональные модели песочных часов отмечают онтогенетические переходы в развитии растений» . Молекулярная биология и эволюция . 33 (5): 1158–1163. DOI : 10.1093 / molbev / msw039 . PMC 4839224 . PMID 26912813 .
- ^ a b Ру, Жюльен; Робинсон-Рехави, Марк (19 декабря 2008 г.). «Ограничения развития на эволюции генома позвоночных» . PLOS Genetics . 4 (12): e1000311. DOI : 10.1371 / journal.pgen.1000311 . ISSN 1553-7404 . PMC 2600815 . PMID 19096706 .
- ^ a b c d Левин, Михал; Анави, Леон; Коул, Элисон Дж .; Зима, Эйтан; Мостова Наталья; Хаир, Салли; Сендерович, Нафтали; Ковалев, Екатерина; Сильвер, Дэвид Х. (31 марта 2016 г.). «Переход к середине развития и эволюция планов тела животных» . Природа . 531 (7596): 637–641. Bibcode : 2016Natur.531..637L . DOI : 10,1038 / природа16994 . ISSN 0028-0836 . PMC 4817236 . PMID 26886793 .
- ^ а б Хейнол, Андреас; Данн, Кейси В. (2016). "Эволюция животных: реальна ли Фила?" . Текущая биология . 26 (10): R424 – R426. DOI : 10.1016 / j.cub.2016.03.058 . PMID 27218852 .