нога членистоногого
Нога членистоногого представляет собой форму суставного придатка членистоногих , обычно используемого для ходьбы . Многие из терминов, используемых для обозначения сегментов ног членистоногих (называемых подомерами ), имеют латинское происхождение и могут быть перепутаны с терминами, обозначающими кости: тазик (что означает бедро , множественное число тазиков ), вертел , бедренная кость (множественное число бедер ), большеберцовая кость (множественное число голеней ), tarsus (множественное число tarsi ), седалищная кость (множественное число седалищных костей ),плюсна , запястье , дактилус (имеется в виду палец ), надколенник (множественное число надколенников ).
Гомологии сегментов ног между группами трудно доказать, и это является источником многих аргументов. Некоторые авторы утверждают, что у самого недавнего общего предка современных членистоногих было до одиннадцати сегментов на ногу [1] , но у современных членистоногих их восемь или меньше. Утверждалось [2] [3] , что наследственная нога не обязательно должна была быть такой сложной, и что другие события, такие как последовательная потеря функции Hox -гена , могли привести к параллельному увеличению сегментов ног.
У членистоногих каждый из сегментов ноги сочленяется со следующим сегментом в шарнирном суставе и может сгибаться только в одной плоскости. Это означает, что требуется большее число сегментов для осуществления тех же движений, которые возможны у позвоночных животных, имеющих вращательные шаровидные суставы в основании передних и задних конечностей. [4]
Двуветвистые и одноветвистые
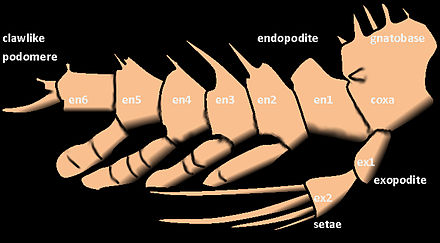
Придатки членистоногих могут быть как двуветвистыми , так и одноветвистыми . Одноветвистая конечность состоит из одной серии сегментов, прикрепленных конец к концу. Однако двуветвистая конечность разветвляется на две, и каждая ветвь состоит из ряда сегментов, прикрепленных конец к концу.
Внешняя ветвь (ветвь) придатков ракообразных известна как экзопод или экзоподит , а внутренняя ветвь известна как эндопод или эндоподит . Другие структуры, кроме двух последних, называются выходами (внешние структуры) и эндитами (внутренние структуры). Экзоподиты легко отличить от выходов по наличию внутренней мускулатуры. Экзоподиты иногда могут отсутствовать у некоторых групп ракообразных ( амфиподы и изоподы ), а у насекомых они полностью отсутствуют. [5]
Ноги насекомых и многоножек одноветвистые. У ракообразных первые усики одноветвистые, а вторые двуветвистые, как и ноги у большинства видов.
Какое-то время обладание одноветвистыми конечностями считалось общим производным признаком , поэтому одноветвистые членистоногие были сгруппированы в таксон под названием Uniramia . В настоящее время считается, что несколько групп членистоногих развили одноветвистые конечности независимо от предков с двуветвистыми конечностями, поэтому этот таксон больше не используется.

{kind=link}
Хелицерата
Ноги паукообразных отличаются от ног насекомых добавлением двух сегментов по обе стороны от большеберцовой кости, надколенника между бедром и большеберцовой костью и плюсневой кости (иногда называемой базитарсусом) между большеберцовой костью и предплюсной (иногда называемой телотарсус), что делает всего семь сегментов.
Лапки пауков имеют когти на конце, а также крючок, который помогает плести паутину. Ноги паука также могут выполнять сенсорные функции, с волосками, которые служат рецепторами прикосновения, а также органом на предплюсне, который служит рецептором влажности, известным как орган предплюсны . [6]
Ситуация идентична у скорпионов , но с добавлением предплюсны за лапкой. Когти скорпиона на самом деле не ноги, а педипальпы , другой вид придатков , которые также встречаются у пауков и специализируются на хищничестве и спаривании.
У Limulus нет ни метатарсусов, ни претарсусов, оставляя по шесть сегментов на ногу.
Ракообразные
Ноги ракообразных примитивно разделены на семь сегментов, которые не соответствуют системе именования, используемой в других группах. Это: тазик, основание, седалищная кость, мерус, запястье, проподус и дактилус. В некоторых группах некоторые сегменты конечностей могут срастаться. Клешня ( чела ) омара или краба образована сочленением дактиля с выростом проподуса. Конечности ракообразных также отличаются двуветвистостью, в то время как у всех других существующих членистоногих конечности одноветвистые.
Многоножки
Многоножки ( многоножки , многоножки и их родственники) имеют семичлениковые ходильные ноги, включающие тазик, вертлуг, предбедренную кость, бедренную кость, большеберцовую кость, предплюсну и коготь предплюсны. Ноги многоножек демонстрируют множество модификаций в разных группах. У всех многоножек первая пара ног преобразована в пару ядовитых клыков, называемых forcipules. У большинства многоножек одна или две пары ходильных ног у взрослых самцов преобразованы в структуры, передающие сперму, называемые гоноподами . У некоторых многоножек первая пара ног у самцов может быть уменьшена до крошечных крючков или обрубков, в то время как у других первая пара может быть увеличена.
Насекомые
Насекомые и их родственники - шестиногие, имеющие шесть ног, соединенных с грудной клеткой , каждая из которых состоит из пяти компонентов. По порядку от тела это тазик, вертел, бедренная кость, большеберцовая кость и предплюсна. Каждый из них представляет собой отдельный сегмент, за исключением лапки, которая может состоять из трех-семи сегментов, каждый из которых называется члеником лапки .
За исключением видов, у которых ноги были утрачены или стали рудиментарными в результате эволюционной адаптации, взрослые насекомые имеют шесть ног, по одной паре, прикрепленных к каждому из трех сегментов грудной клетки. У них есть парные придатки на некоторых других сегментах, в частности, ротовой аппарат , усики и церки , которые произошли от парных ног на каждом сегменте какого-то общего предка .
Однако у некоторых личинок насекомых на брюшных сегментах есть дополнительные ходильные ноги; эти дополнительные ноги называются ложными . Наиболее часто они обнаруживаются на личинках моли и пилильщиков. Проножки не имеют такой же структуры, как ноги современных взрослых насекомых, и было много споров о том, гомологичны ли они им. [7] Текущие данные свидетельствуют о том, что они действительно гомологичны вплоть до очень примитивной стадии своего эмбриологического развития, [8] но их появление у современных насекомых не было гомологичным между чешуекрылыми и симфитами . [9] Такие концепции широко распространены в современных интерпретациях филогенеза. [10]
В целом ноги личинок насекомых, особенно Endopterygota , различаются больше, чем у взрослых особей. Как уже упоминалось, у некоторых есть ложноножки, а также «настоящие» грудные ноги. Некоторые вообще не имеют внешне видимых ног (хотя у них есть внутренние зачатки, которые появляются как взрослые ноги при последнем шелушении ). Примеры включают личинок мух или личинок долгоносиков . Напротив, личинки других жесткокрылых , таких как Scarabaeidae и Dytiscidae , имеют грудные ноги, но не имеют ложноножек. Некоторые насекомые, проявляющие гиперметаморфоз , начинают свой метаморфоз в виде планидий., специализированные, активные личинки с ногами, но они заканчивают свою личиночную стадию как безногие личинки, например Acroceridae .
У Exopterygota ноги личинок имеют тенденцию напоминать ноги взрослых особей в целом, за исключением приспособления к их соответствующему образу жизни. Например, ноги большинства неполовозрелых Ephemeroptera приспособлены к лазанию под подводными камнями и т.п., тогда как у взрослых особей более изящные ноги, которые менее обременительны во время полета. Опять же, детенышей Coccoidea называют «ползунками», и они ползают в поисках подходящего места для кормления, где они оседают и остаются на всю жизнь. У большинства видов их более поздние возрастные стадии не имеют функциональных ног. У Apterygota ноги неполовозрелых особей фактически представляют собой уменьшенные версии ног взрослых особей. [ нужна цитата]
Фундаментальная морфология ног насекомых
Репрезентативная нога насекомого, например, у комнатной мухи или таракана , состоит из следующих частей, расположенных в порядке от наиболее проксимального к наиболее дистальному :
- тазик
- вертел
- бедро
- большеберцовая кость
- предплюсна
- претарсус.
С самой ногой связаны различные склериты вокруг ее основания. Их функции суставные и связаны с тем, как нога прикрепляется к основному экзоскелету насекомого. Такие склериты значительно различаются между неродственными насекомыми. [7]
Кокса
Тазик является проксимальным сегментом и функциональной основой ноги. Он сочленяется с плеврой и связанными с ней склеритами грудного сегмента, а у некоторых видов также сочленяется с краем стернита. Гомология различных базальных склеритов остается предметом споров. Некоторые авторитеты предполагают, что они происходят от предкового subcoxa. У многих видов тазик имеет две доли, где он сочленяется с плеврой. Задняя доля - это мерон , который обычно составляет большую часть тазика. Мерон хорошо развит у Periplaneta, Isoptera, Neuroptera и Lepidoptera.
вертлуг
Вертел сочленяется с тазиком, но обычно жестко прикреплен к бедренной кости. У некоторых насекомых его внешний вид может сбивать с толку; например, у Odonata есть два подсегмента. У паразитических перепончатокрылых основание бедренной кости имеет вид второго вертлуга.
бедренная кость
У большинства насекомых бедренная кость является самой большой частью ноги; это особенно заметно у многих насекомых с сальтаторными ногами, потому что типичный механизм прыжка заключается в выпрямлении сустава между бедренной и большеберцовой костями, а бедренная кость содержит необходимую массивную двуперистую мускулатуру.
большеберцовая кость
Большеберцовая кость — это четвертый отдел типичной ноги насекомого. Как правило, голень насекомого тонкая по сравнению с бедром, но обычно она не меньше, а часто и длиннее. Ближе к дистальному концу обычно имеется большеберцовая шпора, часто две или более. У Apocrita голень передней ноги несет большую апикальную шпору, которая подходит к полукруглой щели в первом сегменте лапки. Щель выстлана гребенчатыми щетинками, и насекомое чистит свои усики, протягивая их.
Тарсус
Предковая лапка была одночлениковой, а у современных Protura , Diplura и некоторых личинок насекомых лапка также одночлениковая. У большинства современных насекомых лапки разделены на подсегменты (членики лапок), обычно около пяти. Фактическое количество варьируется в зависимости от таксона , что может быть полезно для диагностических целей. Например, для Pterogeniidae характерны 5-члениковые передние и средние лапки, но 4-члениковые задние лапки, тогда как Cerylonidae имеют по четыре членика на каждой лапке.
Дистальный сегмент ноги типичного насекомого — претарсус. У Collembola , Protura и многих личинок насекомых претарсус представляет собой единственный коготок. На претарсусе у большинства насекомых есть пара коготков ( ungues , single unguis ). Между когтями претарсус поддерживает срединная пластина unguitractor. Пластинка прикрепляется к аподеме мышцы-сгибателя ног. У Neoptera паремподии представляют собой симметричную пару структур, возникающих на внешней (дистальной) поверхности пластинки унгитрактора между коготками. [11] Он присутствует у многих Hemiptera и почти у всех Heteroptera . [11]Обычно паремподии щетинистые (щетинковидные), но у некоторых видов они мясистые. [12] Иногда паремподии уменьшаются в размерах настолько, что почти исчезают. [13] Над пластиной unguitractor претарсус расширяется вперед в срединную долю, arolium .
Паутинопрядильщики ( Embioptera ) имеют увеличенные базальные членики на каждой из передних ног, содержащие железы , производящие шелк . [14]
Под претарсисом у представителей двукрылых обычно есть парные доли или пульвиллы, что означает «маленькие подушки». Под каждым когтем находится по одной пульвилле. Пульвиллы часто имеют между собой аролиум или, иначе, срединную щетину или эмподий , что означает место встречи пульвилл. На нижней стороне сегментов предплюсны часто имеются пульвилоподобные органы или плантулы.. Аролиум, плантулы и пульвиллы — это адгезивные органы, позволяющие их обладателям взбираться по гладким или крутым поверхностям. Все они являются отростками экзоскелета, а их полости наполнены кровью. Их структуры покрыты трубчатыми щетинковидными волосками, вершины которых смачиваются железистым секретом. Органы приспособлены для плотного прилегания волосков к гладкой поверхности, так что сцепление происходит за счет поверхностных молекулярных сил. [7] [15]
Насекомые контролируют ноготки за счет напряжения мышц длинного сухожилия, «ретрактора ногтя» или «длинного сухожилия». У насекомых, моделирующих передвижение и двигательный контроль, таких как дрозофилы ( Diptera ), саранча ( Acrididae ) или палочники ( Phasmatodea ), длинные сухожилия проходят через предплюсну и большеберцовую кость, прежде чем достичь бедренной кости. Натяжение длинного сухожилия контролируется двумя мышцами, одной в бедре и одной в большеберцовой кости, которые могут работать по-разному в зависимости от того, как нога согнута. Напряжение длинного сухожилия контролирует коготь, но также сгибает предплюсну и, вероятно, влияет на ее жесткость при ходьбе. [16]
Варианты функциональной анатомии ног насекомых
Типичная грудная нога взрослого насекомого приспособлена для бега, а не для копания, прыжков, плавания, хищничества или других подобных действий. Ноги большинства тараканов являются хорошим примером. Однако существует множество специализированных приспособлений, в том числе:
- Передние конечности Gryllotalpidae и некоторых Scarabaeidae приспособлены к рытью нор в земле.
- Передние конечности Mantispidae , Mantodea и Phymatinae приспособлены к захвату и удержанию добычи одним способом, в то время как у Gyrinidae длинные и приспособлены для захвата пищи или добычи совершенно другим способом.
- Передние конечности некоторых бабочек, таких как многие Nymphalidae , редуцированы настолько, что остаются только две пары функциональных ходильных ног.
- У большинства прямокрылых задние ноги скачкообразные ; у них сильно двуперистые мускулистые бедра и прямые длинные голени, приспособленные к прыжкам и, в некоторой степени, к защите ногами. У жуков-блох также есть мощные задние бедра, которые позволяют им эффектно прыгать.
- Другие жуки с эффектно мускулистыми задними бедрами могут вообще не быть скачкообразными, но очень неуклюжими; например, некоторые виды Bruchinae используют свои опухшие задние ноги, чтобы пробиваться из семян растений с твердой оболочкой, таких как Erythrina , в которых они выросли до зрелого возраста.
- Ноги Odonata , стрекоз и стрекоз приспособлены для захвата добычи, которой насекомые питаются во время полета или неподвижно сидя на растении; они почти неспособны использовать их для ходьбы. [7]
- Большинство водных насекомых используют свои ноги только для плавания, хотя многие виды неполовозрелых насекомых плавают другими способами, такими как извиваясь, волнообразно или выбрасывая струи воды.
Эволюция и гомология ног членистоногих
Сегменты эмбрионального тела ( сомиты ) различных таксонов членистоногих разошлись от простого плана тела с множеством сходных придатков, которые серийно гомологичны, к разнообразным планам тела с меньшим количеством сегментов, оснащенных специализированными придатками. [17] Гомологии между ними были обнаружены путем сравнения генов в эволюционной биологии развития . [18]
| Сомит (сегмент тела ) | Трилобит ( Trilobitomorpha ) | Паук ( Хелицерата ) | Многоножка ( Myriapoda ) | Насекомое ( Гексапода ) | Креветки ( Ракообразные ) |
|---|---|---|---|---|---|
| 1 | антенны | хелицеры (челюсти и клыки) | антенны | антенны | 1-е усики |
| 2 | 1-я нога | педипальпы | - | - | 2-е усики |
| 3 | 2 ноги | 1-я нога | челюсти | челюсти | мандибулы (челюсти) |
| 4 | 3 ноги | 2 ноги | 1-я верхняя челюсть | 1-я верхняя челюсть | 1-я верхняя челюсть |
| 5 | 4 ноги | 3 ноги | 2-я верхняя челюсть | 2-я верхняя челюсть | 2-я верхняя челюсть |
| 6 | пятая нога | 4 ноги | коллум (без ножек) | 1-я нога | 1-я нога |
| 7 | 6 ноги | - | 1-я нога | 2 ноги | 2 ноги |
| 8 | 7 ноги | - | 2 ноги | 3 ноги | 3 ноги |
| 9 | 8-я нога | - | 3 ноги | - | 4 ноги |
| 10 | 9 ноги | - | 4 ноги | - | пятая нога |
Классификация модификаций ног
- Ископаемые ноги приспособлены для рытья и рытья нор . Например: медведки имеют роющие передние лапы .
- Сальтаторные ноги приспособлены для прыжков и прыжков. Например: многие прямокрылые имеют скачкообразные задние конечности .
- Нататорские ноги приспособлены для плавания . Например: жуки -вертушки имеют плавающие среднегрудные и заднегрудные ноги .
- Беговые ноги модифицированы для бега . Например: у многих тараканов очень чувствительные беговые ноги .
- Ноги хищника приспособлены для хватания , обычно это приспособление для охоты. Например: богомолы используют свои хищные передние лапы для охоты .
использованная литература
- ^ Кукалова-Пек, Дж. (1992). «Uniramia» не существует - план Pterygota, обнаруженный пермскими Diaphanopterodea из России (Insecta, Paleodictyopteroidea)». Канадский журнал зоологии . 70 (2): 236–255. дои : 10.1139/z92-037 .
- ^ Фрайер, Г. (1996). «Размышления об эволюции членистоногих» . биол. Дж. Линн. соц . 58 (1): 1–55. doi : 10.1111/j.1095-8312.1996.tb01659.x .
- ^ Шрам, ФР и С. Кенеманн (2001). «Генетика развития и эволюция членистоногих: часть I, на ногах». Эволюция и развитие . 3 (5): 343–354. doi : 10.1046/j.1525-142X.2001.01038.x . PMID 11710766 .
- ^ Пэт Уиллмер; Грэм Стоун; Ян Джонстон (12 марта 2009 г.). Экологическая физиология животных . Джон Уайли и сыновья. п. 329. ИСБН 978-1-4443-0922-5.
- ^ Джефф А. Боксхолл и Дамиа Жауме (2009). «Экзоподиты, эпиподиты и жабры ракообразных» (PDF) . Систематика и филогения членистоногих . Музей Tierkunde Дрезден. 67 (2): 229–254. Архивировано (PDF) из оригинала 26 апреля 2019 г .. Проверено 14 января 2012 г. .
- ↑ Пехманн , Матиас (ноябрь 2010 г.). «Механизмы формирования и морфологическое разнообразие придатков пауков и их значение для эволюции пауков» . Строение и развитие членистоногих . 39 (6): 453–467 . Проверено 20 августа 2020 г.
- ^ a b c d Ричардс, О.В.; Дэвис, Р.Г. (1977). Общий учебник энтомологии Иммса: Том 1: Структура, физиология и развитие Том 2: Классификация и биология . Берлин: Спрингер. ISBN 0-412-61390-5.
- ^ Панганибан, Грейс; Надь, Лиза; Кэрролл, Шон Б. «Роль гена Distal-less в развитии и эволюции конечностей насекомых». Текущая биология . 4 (8): 671–675. doi : 10.1016/S0960-9822(00)00151-2 .
- ^ Судзуки, Ю; Палополи, МФ (октябрь 2001 г.). «Эволюция брюшных придатков насекомых: ложноножки гомологичны или конвергентны по признакам?». Дев Гены Эвол . 211 (10): 486–92. doi : 10.1007/s00427-001-0182-3 . PMID 11702198 .
- ^ Галис, Фритсон (1996). «Эволюция насекомых и позвоночных: гомеобоксные гены и гомология». Тенденции в экологии и эволюции . 11 (10): 402–403. doi : 10.1016/0169-5347(96)30038-4 .
- ^ б Фридеманн , Катрин; Спангенберг, Рико; Ёсидзава, Казунор; Бейтель, Рольф Г. (2013). «Эволюция структур прикрепления у самых разнообразных Acercaria (Hexapoda)» (PDF) . Кладистика . 30 : 170–201. doi : 10.1111/cla.12030 . Архивировано из оригинала (PDF) 25 января 2014 года . Проверено 25 января 2014 г.
- ^ Шу, Рэндалл Т. и Слейтер, Джеймс Александр (1995). Настоящие жуки мира (Hemiptera: Heteroptera): классификация и естественная история . Итака, Нью-Йорк: Издательство Корнельского университета. п. 46 . ISBN 978-0-8014-2066-5.
- ^ Гоэль, Южная Каролина (1972). «Заметки о строении пластинки unguitractor у Heteroptera (Hemiptera)». Журнал энтомологии, серия А. 46 (2): 167–173. doi : 10.1111/j.1365-3032.1972.tb00124.x .
- ^ Росс, Эдвард С. (1991). «эмбиоптеры». В Науманн, ID; Карн, ПБ; и другие. (ред.). Насекомые Австралии. Том 1 (2-е изд.). Издательство Мельбурнского университета. стр. 405–409.
- ^ Станислав Н Горб. «Биологические устройства прикрепления: изучение разнообразия природы для биомиметики Phil. Trans. R. Soc. A 2008; 366 (1870): 1557-1574 doi: 10.1098 / rsta.2007.2172 1471-2962
- ^ РАДНИКОВ, Г .; БЕССЛЕР, У. (1 мая 1991 г.). «Функция мышцы, чья аподема проходит через сустав, движимый другими мышцами: почему мышца-ретрактор Unguis у палочников является трехсторонней и не имеет антагониста» . Журнал экспериментальной биологии . 157 (1): 87–99. doi : 10.1242/jeb.157.1.87 . ISSN 0022-0949 .
- ^ Фонд Новартис; Холл, Брайан (2008). Гомология . Джон Уайли. п. 29. ISBN 978-0-470-51566-2.
- ^ Бруска, RC; Бруска, Г.Дж. (1990). Беспозвоночные . Синауэр Ассошиэйтс. п. 669 .
- Морфология членистоногих
- Передвижение животных
- Анатомия паука
- Анатомия трилобита