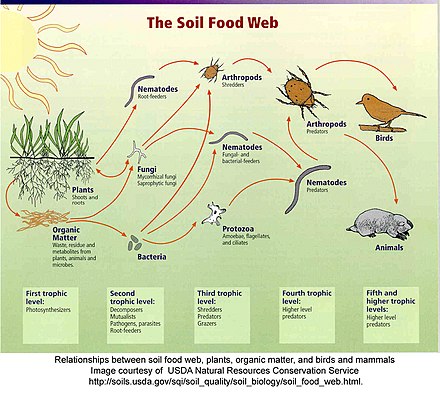
Веб пищи почва это сообщество организмов , живущих всю или часть своей жизни в почве. Он описывает сложную живую систему в почве и то, как она взаимодействует с окружающей средой, растениями и животными.
Пищевые сети описывают передачу энергии между видами в экосистеме . В то время как пищевая цепочка исследует один, линейный, энергетический путь через экосистему, пищевая сеть является более сложной и иллюстрирует все возможные пути. Большая часть этой передаваемой энергии исходит от солнца. Растения используют энергию солнца для преобразования неорганических соединений в богатые энергией органические соединения , превращая углекислый газ и минералы в растительный материал путем фотосинтеза . Цветы растений источают богатый энергией нектар над землей, а корни растений источают кислоты, сахара и эктоферменты в ризосферу , регулируя pH.и подпитка пищевой сети под землей. [2] [3] [4]
Растения называют автотрофами, потому что они вырабатывают собственную энергию; их также называют производителями, потому что они производят энергию, доступную для употребления в пищу другими организмами. Гетеротрофы - это потребители, которые не могут самостоятельно готовить пищу. Чтобы получить энергию, они едят растения или других гетеротрофов.
Надземные пищевые сети [ править ]
В наземных пищевых сетях энергия перемещается от производителей (растений) к основным потребителям ( травоядные животные ), а затем к вторичным потребителям (хищники). Фраза « трофический уровень» относится к различным уровням или этапам энергетического пути. Другими словами, продуценты, потребители и деструкторы являются основными трофическими уровнями. Эта цепочка передачи энергии от одного вида к другому может продолжаться еще несколько раз, но в конце концов заканчивается. В конце пищевой цепочки такие разложители, как бактерии и грибы, расщепляют мертвые растения и животные на простые питательные вещества.
Методология [ править ]
Природа почвы затрудняет прямое наблюдение за пищевыми цепями. Поскольку почвенные организмы имеют размер от менее 0,1 мм (нематоды) до более 2 мм (дождевые черви), существует множество различных способов их извлечения. Образцы почвы часто берут с помощью металлического керна. Крупную макрофауну, такую как дождевые черви и личинки насекомых, можно удалить вручную, но это невозможно для более мелких нематод и почвенных членистоногих. Большинство методов извлечения мелких организмов являются динамическими; они зависят от способности организмов выходить из почвы. Например, воронка Берлезе , используемая для сбора мелких членистоногих., создает градиент света / тепла в образце почвы. По мере того, как микроартроподы движутся вниз, прочь от света и тепла, они падают через воронку в пробирку для сбора. Похожий метод - воронка Бермана - используется для нематод. Однако воронка Баермана мокрая (в то время как воронка Берлезе сухая) и не зависит от градиента света / тепла. Нематоды выходят из почвы на дно воронки, потому что при движении они плотнее воды и не могут плавать. Сообщества почвенных микробов характеризуются по-разному. Активность микробов можно измерить по их дыханию и выделению углекислого газа. Клеточные компоненты микробов могут быть извлечены из почвы и генетически профилированы, или микробная биомасса может быть рассчитана путем взвешивания почвы до и после фумигации.
Типы пищевых сетей [ править ]

{kind=link}
Существует три различных типа представлений пищевой сети: топологические (или традиционные) пищевые сети, сети потока и сети взаимодействия. Эти сети могут описывать системы как над землей, так и под землей.
Топологические сети [ править ]
Ранние пищевые сети были топологическими; они были описательными и давали неколичественную картину потребителей, ресурсов и связей между ними. Pimm et al. (1991) описали эти сети как карту того, какие организмы в сообществе питаются другими видами. Самая ранняя топологическая пищевая сеть, созданная в 1912 году, была посвящена исследованию хищников и паразитов хлопкового долгоносика (обзор Пимма и др., 1991). Исследователи проанализировали и сравнили топологические сети между экосистемами, измеряя длину цепочки взаимодействия и возможности подключения. [5]Одна из проблем, с которыми сталкивается при стандартизации таких измерений, заключается в том, что часто бывает слишком много видов для каждого, чтобы иметь отдельный ящик. В зависимости от автора количество видов, агрегированных или разделенных на функциональные группы, может быть разным. [6] Авторы могут даже исключить некоторые организмы. По соглашению, мертвый материал, возвращающийся к детриту, не показан, поскольку это усложняет фигуру, но это учитывается при любых расчетах. [6]
Сети потока [ править ]
Миоз строится на взаимосвязанных пищевых цепочках, добавляя количественную информацию о перемещении углерода или других питательных веществ от производителей к потребителям. Hunt et al. (1987) опубликовали первую сеть потоков для почвы, описывающую низкотравную прерию в Колорадо, США. Авторы оценили скорость переноса азота через почвенную пищевую сеть и рассчитали скорость минерализации азота для ряда почвенных организмов. В другом знаменательном исследовании исследователи из экспериментальной фермы Ловинхув в Нидерландах изучили поток углерода и проиллюстрировали скорость переноса стрелками разной толщины. [7]
Чтобы создать потоковую сеть, сначала создается топологическая сеть. После определения членов сети рассчитывается биомасса каждой функциональной группы, обычно в кг углерода на гектар . Чтобы рассчитать нормы кормления, исследователи предполагают, что популяция функциональной группы находится в равновесии. При равновесии воспроизводство группы уравновешивает скорость, с которой члены теряются в результате естественной смерти и хищничества [8]. Когда скорость кормления известна, можно рассчитать эффективность, с которой питательные вещества превращаются в биомассу организма. Эта энергия, запасенная в организме, представляет собой количество, доступное для передачи на следующий трофический уровень.
После создания первых сетей почвенного потока исследователи обнаружили, что питательные вещества и энергия перетекают из более низких ресурсов на более высокие трофические уровни по трем основным каналам. [7] [8] Бактериальные и грибковые каналы имели самый большой поток энергии, в то время как канал травоядных, в котором организмы напрямую потребляли корни растений, был меньше. В настоящее время широко признано, что бактерии и грибки имеют решающее значение для разложения углерода и азота и играют важную роль как в углеродном, так и в азотном циклах .
Интернет взаимодействия [ править ]
Сеть взаимодействия, показанная справа вверху [9] , похожа на топологическую сеть, но вместо того, чтобы показывать движение энергии или материалов, стрелки показывают, как одна группа влияет на другую. В моделях пищевой сети взаимодействия каждая ссылка имеет два прямых воздействия: ресурс на потребителя и один на потребителя на ресурс. [10] Влияние ресурса на потребителя положительное (потребитель получает возможность есть), а влияние потребителя на ресурс отрицательное (он съедается). Эти прямые, трофические эффекты могут приводить к косвенным эффектам. Косвенные эффекты, представленные пунктирными линиями, показывают влияние одного элемента на другой, с которым он напрямую не связан. [10]Например, в простой сети взаимодействия ниже, когда хищник ест корневое травоядное животное, у растения, съеденного травоядным, может увеличиться биомасса. Тогда мы бы сказали, что хищник оказывает косвенное благотворное влияние на корни растений.
Контроль пищевой сети [ править ]
Эффекты снизу вверх [ править ]
Эффект снизу вверх возникает, когда плотность ресурса влияет на плотность его потребителя. [11] Например, на рисунке выше увеличение плотности корней вызывает увеличение плотности травоядных, что вызывает соответствующее увеличение плотности хищников. Корреляция в изобилии или биомассе между потребителями и их ресурсами свидетельствует о восходящем контроле. [11] Часто упоминаемый пример восходящего эффекта - это взаимосвязь между травоядными животными и первичной продуктивностью растений. В наземных экосистемах биомасса травоядных и детритофаговувеличивается с первичной продуктивностью. Увеличение первичной продуктивности приведет к большему притоку опада из листьев в почвенную экосистему, что предоставит больше ресурсов для роста популяций бактерий и грибов. Увеличение количества микробов приведет к увеличению количества питающихся бактериями и грибами нематод , которых поедают клещи и другие хищные нематоды. Таким образом, вся пищевая сеть раздувается по мере того, как к базе добавляются новые ресурсы. [11] Когда экологи используют термин «восходящий контроль», они указывают, что биомасса, численность или разнообразие более высоких трофических уровней зависят от ресурсов более низких трофических уровней. [10]
Эффекты сверху вниз [ править ]
Идеи о контроле сверху вниз оценить гораздо труднее. Эффекты сверху вниз возникают, когда плотность населения потребителя влияет на плотность его ресурса; [10] например, хищник влияет на плотность своей добычи. Следовательно, нисходящий контроль относится к ситуациям, когда численность, разнообразие или биомасса более низких трофических уровней зависит от воздействия потребителей на более высоких трофических уровнях. [10] трофический каскад представляет собой тип взаимодействия сверху вниз , который описывает косвенное воздействие хищников. В трофическом каскаде хищники вызывают эффекты, которые спускаются вниз по пищевой цепи и влияют на биомассу организмов на расстоянии как минимум двух звеньев. [10]
Важность трофических каскадов и нисходящего контроля в наземных экосистемах активно обсуждается в экологии (обзор в Шурин и др., 2006), и вопрос о том, возникают ли трофические каскады в почвах, не менее сложен [12]. Трофические каскады действительно встречаются в обеих бактериальные и грибковые энергетические каналы. [13] [14] [15] Однако каскады могут быть нечастыми, потому что многие другие исследования не показывают влияния хищников сверху вниз. [16] [17] В исследовании Микола и Сетяля микробы, поедаемые нематодами, росли быстрее, если их часто пасли. Этот компенсаторный ростзамедлился, когда были удалены нематоды, питающиеся микробами. Следовательно, хотя высшие хищники сократили количество питающихся нематод микробами, общего изменения микробной биомассы не произошло.
Помимо эффекта выпаса, еще одним препятствием на пути к контролю сверху вниз в почвенных экосистемах является широко распространенная всеядность, которая, увеличивая количество трофических взаимодействий, ослабляет воздействие сверху. Почвенная среда также представляет собой матрицу различных температур, влажности и уровней питательных веществ, и многие организмы способны переходить в спящий режим, чтобы противостоять трудным временам. В зависимости от условий хищники могут быть отделены от своей потенциальной жертвы непреодолимым пространством и временем.
Любые возникающие нисходящие эффекты будут ограничены по силе, поскольку почвенные пищевые сети контролируются донорами. Контроль доноров означает, что потребители практически не влияют на обновление или ввод своих ресурсов. [10] Например, наземные травоядные животные могут перевыпасывать территорию и уменьшать популяцию травы, но разлагатели не могут напрямую влиять на скорость падения опада. Они могут лишь косвенно влиять на скорость поступления в свою систему питательных веществ, рециркулируя питательные вещества, которые, помогая растениям расти, в конечном итоге создают больше опада и мусора. [18] Однако, если бы вся трофическая сеть почвы находилась под полным контролем доноров, бактериоядные и грибковые животные никогда бы не повлияли на потребляемые ими бактерии и грибки.
Хотя восходящие эффекты, несомненно, важны, многие экологи почвы подозревают, что нисходящие эффекты также иногда значительны. Некоторые хищники или паразиты при добавлении в почву могут оказывать сильное воздействие на корневых травоядных и тем самым косвенно влиять на приспособленность растений. Например, в пищевой цепи прибрежных кустарников корни кустарникового люпина поедали аборигенные энтомопатогенные нематоды , Heterorhabditis marelatus , паразитированные гусеницы-призраки и гусеницы-призраки. Наличие H. marelatusкоррелирует с меньшим количеством гусениц и более здоровыми растениями. Кроме того, исследователи наблюдали высокую смертность люпина кустарного в отсутствие энтомопатогенных нематод. Эти результаты предполагают, что нематода, как естественный враг гусеницы призрачной моли, защищает растение от повреждений. Авторы даже предположили, что взаимодействие было достаточно сильным, чтобы повлиять на динамику популяций люпина кустового; [19] это было подтверждено в более поздних экспериментальных работах с естественно растущими популяциями кустарникового люпина. [20]
Контроль сверху вниз находит применение в сельском хозяйстве и является принципом биологического контроля , идеей о том, что растения могут получить выгоду от применения врагов их травоядных. В то время как осы и божьи коровки обычно связаны с биологической борьбой, паразитические нематоды и хищные клещи также добавляются в почву для подавления популяций вредителей и сохранения сельскохозяйственных культур. Для эффективного использования таких агентов биологической борьбы важно знание местной пищевой сети почвы.
Матричные модели сообщества [ править ]
Сообщество матрица модель представляет собой тип веб - взаимодействия , который использует дифференциальные уравнения для описания каждой ссылки в топологической сети. С помощью уравнений Лотки – Вольтерры , которые описывают взаимодействия хищников и жертв, и данных по энергетике пищевой сети, таких как биомасса и скорость кормления, рассчитывается сила взаимодействий между группами. [21] Матричные модели сообщества также могут показать, как небольшие изменения влияют на общую стабильность сети.
Устойчивость пищевых сетей [ править ]
Математическое моделирование пищевых сетей подняло вопрос о том, какие пищевые сети более устойчивы: сложные или простые. До последнего десятилетия считалось, что почвенные пищевые сети относительно просты, с низкой степенью связи и всеядностью. [12] Эти идеи возникли из математических моделей Мэя, которые предсказывали, что сложность дестабилизирует пищевые сети. Мэй использовал матрицы сообществ, в которых виды были случайным образом связаны с силой случайного взаимодействия, чтобы показать, что локальная стабильность снижается со сложностью (измеряемой как взаимосвязь), разнообразием и средней силой взаимодействия между видами. [22]
Использование таких случайных матриц сообществ вызвало много критики. В других областях экологии было обнаружено, что пищевые сети, использованные для создания этих моделей, были сильно упрощены [23] и не отражали сложность реальных экосистем. Также стало ясно, что почвенные пищевые сети не соответствовали этим предсказаниям. Экологи почв обнаружили, что всеядность в пищевых сетях была обычным явлением [24], и что пищевые цепи могли быть длинными и сложными [8] и при этом оставаться устойчивыми к нарушениям в результате сушки, замораживания и окуривания. [12]
Но почему сложные пищевые сети более устойчивы? Многие из препятствий на пути нисходящих трофических каскадов также способствуют стабильности. Сложные пищевые сети могут быть более стабильными, если силы взаимодействия слабые [22], а почвенные пищевые сети, по-видимому, состоят из множества слабых взаимодействий и нескольких сильных. [21] Пищевые сети, контролируемые донорами, могут быть по своей природе более стабильными, потому что первичным потребителям сложно перенапрягать свои ресурсы. [25] Структура почвы также действует как буфер, разделяя организмы и предотвращая сильные взаимодействия. [12] Многие почвенные организмы, например бактерии, могут оставаться в состоянии покоя в трудные времена и быстро размножаться при улучшении условий, что делает их устойчивыми к нарушениям.
Устойчивость системы снижается за счет использования азотсодержащих неорганических и органических удобрений, вызывающих закисление почвы .
Взаимодействия, не включенные в пищевые сети [ править ]
Несмотря на их сложность, некоторые взаимодействия между видами в почве нелегко классифицировать с помощью пищевых сетей. Трансформеры мусора, мутуалисты и инженеры экосистемы - все они оказывают сильное влияние на свои сообщества, которое нельзя охарактеризовать как восходящее или нисходящее.
Трансформаторы подстилки, такие как равноногие , поедают мертвые растения и выделяют фекальные гранулы. Хотя на первый взгляд это может показаться не впечатляющим, фекальные гранулы более влажные и содержат больше питательных веществ, чем окружающая почва, что способствует колонизации бактериями и грибами. Разложение фекальных гранул микробами увеличивает их питательную ценность, и изопод может повторно проглатывать гранулы. Когда равноногие моллюски потребляют подстилку с низким содержанием питательных веществ, микробы обогащают ее для них, и изоподы, которым не дают поедать собственные фекалии, могут погибнуть. [26] Эти взаимные отношения были названы «внешним рубцом», аналогично взаимным отношениям между бактериями и коровами. В то время как бактериальные симбионты коров живут внутри рубца их желудка равноногие зависят от микробов вне их тела.
Инженеры экосистем, такие как дождевые черви, изменяют свою среду обитания и создают среду обитания для других более мелких организмов. Дождевые черви также стимулируют микробную активность, увеличивая аэрацию и влажность почвы и перенося мусор в землю, где он становится доступным для другой почвенной фауны. [12] Грибы создают пищевую нишу для других организмов, обогащая крайне дефицитную пищу - мертвую древесину. [27] Это позволяет ксилофагам развиваться и, в свою очередь, воздействовать на мертвую древесину, способствуя разложению древесины и круговороту питательных веществ в лесной подстилке. [28]В надземных и водных пищевых сетях в литературе предполагается, что наиболее важными взаимодействиями являются конкуренция и хищничество. Хотя почвенные пищевые сети хорошо подходят для такого рода взаимодействий, будущие исследования должны включать более сложные взаимодействия, такие как мутуализм и изменение среды обитания.
Хотя они не могут охарактеризовать все взаимодействия, почвенные пищевые сети остаются полезным инструментом для описания экосистем. Взаимодействие между видами в почве и их влияние на разложение продолжают хорошо изучаться. Однако многое остается неизвестным о стабильности почвенных пищевых сетей и о том, как пищевые сети меняются с течением времени. [12] Эти знания имеют решающее значение для понимания того, как пищевые сети влияют на такие важные качества, как плодородие почвы .
См. Также [ править ]
- Биология почвы
- Экология почвы
- Функции почвы
- Почвенная жизнь
- Трофический каскад
- Пищевой сети
- Энтомопатогенная нематода
Ссылки [ править ]
- ^ "Фотогалерея грунтовки биологии" . Служба охраны природных ресурсов - Почвы . Общество охраны почв и водных ресурсов Министерства сельского хозяйства США . Проверено 14 августа 2016 года .
- ^ Маршнер, Хорст (1995). Минеральное питание высших растений . ISBN 978-0124735439.
- ^ Уокер, TS; Байс, HP; Grotewold, E .; Виванко, Дж. М. (2003). «Корневая экссудация и биология ризосферы» . Физиология растений . 132 (1): 44–51. DOI : 10.1104 / pp.102.019661 . PMC 1540314 . PMID 12746510 .
- ^ Мощность, Майкл Л. (2010). Энн М. Берроуз; Линн Т. Нэш (ред.). Эволюция экссудативной болезни у приматов / Проблемы питания и пищеварения для приматов, питающихся жевательной резинкой . Springer. п. 28. ISBN 9781441966612. Проверено 2 октября 2012 года .
- ^ Pimm SL, Lawton JH & Cohen JE (1991), "Модели пищевой сети и их последствия", Nature , 350 (6320): 669–674, Bibcode : 1991Natur.350..669P , doi : 10.1038 / 350669a0 , S2CID 4267587
- ^ a b de Ruiter PC; AM Neutel; Дж. К. Мур (1996), «Энергетика и стабильность подземных пищевых сетей», в G. Polis; KO Winemiller (ред.), Пищевые сети: интеграция моделей и динамики , Chapman & Hall
- ^ a b Brussaard, LJ, A. van Veen, MJ Kooistra и G. Lebbink (1988), «Голландская программа по экологии почв в системах земледелия I. Цели, подход и предварительные результаты», Ecological Bulletins , 39 : 35–40CS1 maint: несколько имен: список авторов ( ссылка )
- ^ a b c Hunt, HW, DC Coleman, ER Ingham, RE Ingham, ET Elliott, JC Moore, SL Rose, CPP Reid и CR Morley (1987), "Обломочная пищевая сеть в низкотравных прериях", Биология и плодородие Почв , 3 : 57–68CS1 maint: multiple names: authors list (link)
- ^ USDA -NRCS, 2004, «Пищевая сеть почвы» в The Soil Biology Primer . URL, доступ к которому осуществлялся 04-11-2006 г.
- ^ a b c d e f g Стилинг, П. (1999), Экология, теории и приложения. Третий Edtn , Prentice Hall. Нью-Джерси США.
- ^ a b c Шурин, Дж. Б., Д. С. Грунер и Х. Хиллебранд (2006), «Все влажные или высохшие? Реальные различия между водными и наземными пищевыми цепями», Proceedings of the Royal Society , 273 (1582): 1–9 , DOI : 10.1098 / rspb.2005.3377 , PMC 1560001 , PMID 16519227 CS1 maint: multiple names: authors list (link)
- ^ a b c d e f Уордл, Д. А. (2002), Сообщества и экосистемы: связь надземных и подземных компонентов Монографии по популяционной биологии, 31 , Princeton University Press. Нью-Джерси
- ^ Сантос, П. Ф., Дж. Филлипс и В. Г. Уитфорд (1981), «Роль клещей и нематод на ранних стадиях разложения подстилки в пустыне», Экология (Вашингтон, округ Колумбия) , 62 (3): 664–669, DOI : 10,2307 / 1937734 , JSTOR 1937733 CS1 maint: multiple names: authors list (link)
- ^ Allen-Morley, CR & DC Coleman (1989), "Reliance биоты почвы в различных пищевых цепочках до замораживания возмущения", экология , 70 (4): 1127-1141, DOI : 10,2307 / 1941381 , JSTOR 1941381
- ^ Katarina Хедлунд и Мария Шегрена Öhrn (2000), "Tritrophic взаимодействия в обществе почвы увеличивают темпы разложения" , Oikos , 88 (3): 585-591, DOI : 10,1034 / j.1600-0706.2000.880315.x , архивируются из оригинал от 05.01.2013
- ^ Микола J. & H. Setälä (1998), "Нет доказательств трофических каскадов в экспериментальной микробной основе трофической", экология , 79 : 153-164, DOI : 10,1890 / 0012-9658 (1998) 079 [0153: NEOTCI] 2.0.CO; 2
- ^ Лааксо Дж. И Х. Сетэля (1999), «Популяционные и экосистемные эффекты хищничества на питающихся микробами нематод» , Oecologia , 120 (2): 279–286, Bibcode : 1999Oecol.120..279L , doi : 10.1007 / s004420050859 , PMID 28308090 , S2CID 21444364
- ^ Мур, JC, K. McCann, H. Setälä, и PC de Ruiter (2003), «Сверху вниз - снизу вверх: регулирует ли хищничество в ризосфере надземную динамику?», Ecology , 84 (4): 846–857, DOI : 10.1890 / 0012-9658 (2003) 084 [0846]: TIBDPI 2.0.CO; 2CS1 maint: multiple names: authors list (link)
- ^ Стронг, Д. Р., Х. К. Кайя, А. В. Уиппл, А. Л., Чайлд, С. Крейг, М. Бондонно, К. Дайер и Дж. Л. Марон (1996), "Энтомопатогенные нематоды: естественные враги корнеплодных гусениц на кустарниковом люпине", Oecologia (Берлин) , 108 (1): 167–173, Bibcode : 1996Oecol.108..167S , doi : 10.1007 / BF00333228 , PMID 28307747 , S2CID 35889439 CS1 maint: multiple names: authors list (link)
- ^ Evan L. Preisser & Donald R. Strong (2004), "Климат влияет на контроль хищника травоядным вспышки", американский натуралист , 163 (5): 754-762, DOI : 10,1086 / 383620 , PMID 15122492 , S2CID 1328187
- ^ a b de Ruiter, PC, Neutel, A.-M., & Moore, JC; Neutel; Moore (1995), "Энергетика, закономерности сильных взаимодействий и стабильности в реальных экосистемах", Science , 269 (5228): 1257-1260, Bibcode : 1995Sci ... 269.1257D , дой : 10.1126 / science.269.5228.1257 , PMID 17732112 , S2CID 30877530 CS1 maint: multiple names: authors list (link)
- ^ а б Май, Р. М. (1973), «Стабильность и сложность в модельных экосистемах», Монографии по популяционной биологии , Принстон: Princeton University Press. Нью-Джерси, 6 : 1–235, PMID 4723571
- ^ Polis, GA (1991), "Комплексные трофические взаимодействия в пустынях: эмпирический критик теории трофической", американский натуралист , 138 : 123-155, DOI : 10,1086 / 285208 , S2CID 84458020
- ^ Walter, DEDT Kaplan & TA Permar (1991), "Недостающие звенья: обзор методов , используемых для оценки трофических связей в пищевых цепочках" , сельское хозяйство, экосистемы и окружающая среда , 34 : 399-405, DOI : 10.1016 / 0167-8809 ( 91) 90123-Ф
- ^ Де Анжелис, DL (1992), Динамика круговорота питательных веществ и пищевых сетей , Чепмен и Холл. Лондон. Англия, ISBN 978-0-12-088458-2
- ^ Hassall, M., SP Rushton (1982), "Роль копрофагии в стратегиях питания наземных равноногих", Oecologia , 53 (3): 374–381, Bibcode : 1982Oecol..53..374H , doi : 10.1007 / BF00389017 , PMID 28311744 , S2CID 38644608 CS1 maint: multiple names: authors list (link)
- ^ Филипьяк, Михал; Собчик, Лукаш; Вайнер, январь (2016-04-09). «Грибковое преобразование пней в подходящий ресурс для жуков-ксилофагов через изменения в соотношении элементов» . Насекомые . 7 (2): 13. DOI : 10,3390 / insects7020013 . PMC 4931425 .
- ^ Филипьяк, Михал; Вайнер, январь (2016-09-01). «Динамика питания в процессе развития жуков-ксилофагов, связанная с изменением стехиометрии 11 элементов» . Физиологическая энтомология . 42 : 73–84. DOI : 10.1111 / phen.12168 . ISSN 1365-3032 .
Внешние ссылки [ править ]
- Лаборатория Soil Foodweb в Канаде
- Soil Foodweb Inc международная сеть лабораторий по анализу почвы и образовательных учреждений
| vтеПочвоведение | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Основные поля |
| |||||||||||
| Почвенные темы |
| |||||||||||
| Тип почвы |
| |||||||||||
| Приложения |
| |||||||||||
| Связанные поля |
| |||||||||||
| Общества, инициативы |
| |||||||||||
| Научные журналы |
| |||||||||||
| Смотрите также |
| |||||||||||
| ||||||||||||