| Yersinia pestis | |
|---|---|
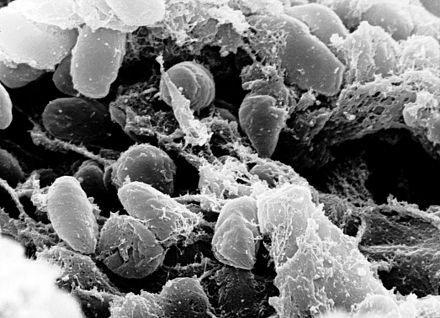 | |
| Сканирующие электронный микроснимок , изображающий массу чумной палочка бактерии в передней кишке инфицированной блохи | |
| Научная классификация | |
| Домен: | Бактерии |
| Тип: | Протеобактерии |
| Учебный класс: | Гаммапротеобактерии |
| Заказ: | Enterobacterales |
| Семья: | Yersiniaceae |
| Род: | Иерсиния |
| Разновидность: | Y. pestis |
| Биномиальное имя | |
| Yersinia pestis (Леманн и Нойман, 1896 г.) ван Логем, 1944 г. | |
| Синонимы | |
| |
{kind=link}
Чумная палочка (ранее Pasteurella Pestis ) представляет собой грамотрицательные , неподвижные , палочковидные , коккобактерия бактерии, без спор. Это факультативный анаэробный организм, который может инфицировать людей через блоху восточных крыс ( Xenopsylla cheopis ). [1] Это вызывает чуму , которая принимает три основные формы: легочную , септическую и бубонную . Могут существовать доказательства того, что Y. pestis возник в Европе в культуре Cucuteni-Trypillia.а не в Азии, как принято считать. [2]
Y. pestis был открыт в 1894 году Александром Йерсеном , швейцарским / французским врачом и бактериологом из Института Пастера , во время эпидемии чумы в Гонконге . [3] Йерсен был членом школы Пастера . Китасато Шибасабуро , японский бактериолог, практикующий методологию Коха , в то время также занимался поиском возбудителя чумы. [4] Однако на самом деле Йерсин связал чуму с Y. pestis . Ранее назывался Pasteurella pestis.В 1944 году этот организм был переименован в Yersinia pestis .
Ежегодно Всемирная организация здравоохранения по- прежнему сообщает о тысячах случаев чумы , хотя при правильном лечении антибиотиками прогноз для жертв сейчас намного лучше. Во время войны во Вьетнаме число случаев заболевания в Азии увеличилось в пять-шесть раз , возможно, из-за нарушения экосистем и более тесной близости между людьми и животными. В настоящее время чума обычно встречается в Африке к югу от Сахары и на Мадагаскаре, в районах, на которые в настоящее время приходится более 95% зарегистрированных случаев. Чума также оказывает пагубное воздействие на млекопитающих, кроме человека; [5] в Соединенных Штатах к ним относятся чернохвостая луговая собачка и находящийся под угрозой исчезновения черноногий хорек..
Общие характеристики [ править ]
Y. pestis - неподвижная, факультативная анаэробная бактерия в форме палочки с биполярным окрашиванием (придавая ей вид английской булавки ), которая образует слой антифагоцитарной слизи. [6] Подобно другим видам Yersinia , он дает отрицательный результат на уреазу , ферментацию лактозы и индол . [7] Его ближайшим родственником является возбудитель желудочно-кишечного тракта Yersinia pseudotuberculosis , а более отдаленно - Yersinia enterocolitica .
Геном [ править ]
Полная геномная последовательность доступна для двух из трех подвидов Y. pestis : штамма KIM (биовара Y. p. Medievalis ) [8] и штамма CO92 (биовара Y. p. Orientalis , полученного из клинического изолята в Соединенные Штаты). [9] По состоянию на 2006 год, геномная последовательность штамма биовара Antiqua была недавно завершена. [10] Подобно другим патогенным штаммам, существуют признаки мутации потери функции. Длина хромосомы штамма KIM составляет 4600755 пар оснований; длина хромосомы штамма CO92 составляет 4 653 728 пар оснований. Подобно Y. pseudotuberculosis иY. enterocolitica , Y. pestis является хозяином плазмиды pCD1. Он также содержит две другие плазмиды, pPCP1 (также называемые pPla или pPst) и pMT1 (также называемые pFra), которые не переносятся другими видами Yersinia . pFra кодирует фосфолипазу D, которая важна для способности Y. pestis передаваться блохами. [11] pPla кодирует протеазу Pla, которая активирует плазмин у человека-хозяина и является очень важным фактором вирулентности при легочной чуме. [12] Вместе эти плазмиды и остров патогенностиназываемый HPI, кодирует несколько белков, вызывающих патогенез, которым известен Y. pestis . Среди прочего, эти факторы вирулентности необходимы для бактериальной адгезии и инъекции белков в клетку-хозяина, вторжения бактерий в клетку-хозяина (через систему секреции типа III ), а также приобретения и связывания железа, полученного из эритроцитов ( по сидерофорам ). Считается, что Y. pestis произошел от Y. pseudotuberculosis и отличается только наличием плазмид специфической вирулентности.
Комплексный и сравнительный протеомный анализ штамма Y. pestis KIM был проведен в 2006 году. [13] Анализ был сосредоточен на переходе к условиям роста, имитирующим рост в клетках-хозяевах.
Малая некодирующая РНК [ править ]
Было обнаружено, что многочисленные бактериальные малые некодирующие РНК выполняют регуляторные функции. Некоторые могут регулировать гены вирулентности. Около 63 новых предполагаемых мРНК были идентифицированы посредством глубокого секвенирования мРНК-ома Y. pestis . Среди них был Yersinia- специфический (также присутствующий у Y. pseudotuberculosis и Y. enterocolitica ) Ysr141 ( малая РНК Yersinia 141). Было показано, что мРНК Ysr141 регулирует синтез эффекторного белка системы секреции типа III (T3SS) YopJ. [14] Yop-Ysc T3SS является критическим компонентом вирулентности для видов Yersinia . [15] Многие новые мРНК были идентифицированы из Y. pestis, выращенного in vitro, и в инфицированных легких мышей, что позволяет предположить, что они играют роль в бактериальной физиологии или патогенезе. Среди них предсказано, что sR035 спаривается с SD-областью и сайтом инициации транскрипции термочувствительного регулятора ymoA, а sR084, как предсказано, спаривается с мехом , регулятором захвата железа . [16]
Патогенез и иммунитет [ править ]
В городском и лесном циклах Y. pestis чаще всего распространяется между грызунами и блохами. В лесном цикле грызун является диким, но в городском цикле грызун - это прежде всего коричневая крыса ( Rattus norvegicus ). Кроме того, Y. pestis может распространяться из городской среды и обратно. Передача людям обычно происходит через укусы инфицированных блох. Если заболевание перешло в легочную форму, люди могут передавать бактерии другим путем кашля, рвоты и, возможно, чихания.
В резервуаре хозяев [ править ]
Несколько видов грызунов служат основным резервуаром Y. pestis в окружающей среде. В степи , то естественный водоем , как полагают, главным образом сурка . Считается, что на западе США несколько видов грызунов поддерживают Y. pestis . Однако ожидаемой динамики заболевания ни у одного грызуна не обнаружено. Известно, что несколько видов грызунов обладают переменной резистентностью, что может привести к бессимптомному статусу носителя . [17] Свидетельства указывают на то, что блохи от других млекопитающих играют роль в вспышках чумы среди людей. [18]
Отсутствие знаний о динамике распространения чумы у млекопитающих также характерно для восприимчивых грызунов, таких как чернохвостая луговая собачка ( Cynomys ludovicianus ), у которой чума может вызывать коллапс колонии, что приводит к сильному воздействию на пищевые сети прерий. [19] Однако динамика передачи у луговых собачек не соответствует динамике блокированных блох; Вместо этого могут быть важны туши, незаблокированные блохи или другой переносчик. [20]
В других регионах мира резервуар инфекции четко не определен, что затрудняет программы профилактики и раннего предупреждения. Один такой пример был замечен во время вспышки болезни в Алжире в 2003 году . [21]
Вектор [ править ]
Передача Y. pestis блохами хорошо изучена. [22] Первоначальное приобретение Y. Pestis по вектору происходит во время кормления на зараженном животном. Затем несколько белков способствуют поддержанию бактерий в пищеварительном тракте блох, среди них система хранения гемина и мышиный токсин Yersinia (Ymt). Хотя Ymt очень токсичен для грызунов и когда-то считалось, что он производится для повторного заражения новых хозяев, он важен для выживания Y. pestis у блох. [11]
Система хранения гемина играет важную роль в передаче Y. pestis обратно млекопитающему-хозяину. [23] Находясь в векторе насекомых, белки, кодируемые генетическими локусами системы хранения гемина, индуцируют образование биопленки в преджелудке , клапане, соединяющем среднюю кишку с пищеводом . [24] [25] Наличие этой биопленки, вероятно, необходимо для стабильной инфекции блох. [26] Агрегация в биопленке препятствует питанию, поскольку образуется масса свернувшейся крови и бактерий (названная «блоком Бакота» по имени энтомолога А.В. Бакота , первого, кто описал это явление).[27] Передача Y. pestis происходит во время тщетных попыток блох прокормиться. Проглоченная кровь закачивается в пищевод, где она вытесняет бактерии, застрявшие в преджелудках, которые срыгивают обратно в кровеносную систему хозяина. [27]
У людей и других восприимчивых хозяев [ править ]
Патогенез из - за Y. Pestis инфекции хостов млекопитающих обусловлено несколькими факторами, в том числе способности этих бактерий , чтобы подавить и избежать нормальной системы иммунной реакции , такие как фагоцитоз и антитела производства. Укусы блох позволяют бактериям проходить через кожный барьер. Y. pestis экспрессирует активатор плазмина , который является важным фактором вирулентности для легочной чумы и может разрушаться на сгустках крови для облегчения систематической инвазии. [12] Многие факторы вирулентности бактерий имеют антифагоцитарную природу. Два важных антифагоцитарных антигена, названные F1 (фракция 1) и V или LcrV , оба важны для вирулентности . [6] Эти антигены продуцируются бактериями при нормальной температуре человеческого тела. Кроме того, Y. pestis выживает и продуцирует антигены F1 и V, находясь в белых кровяных тельцах, таких как моноциты , но не в нейтрофилах . Естественный или индуцированный иммунитет достигается за счет продукции специфических опсонических антител против антигенов F1 и V; антитела против F1 и V вызывают фагоцитоз нейтрофилов. [28]
Кроме того, система секреции типа III (T3SS) позволяет Y. pestis вводить белки в макрофаги и другие иммунные клетки. Эти инъецированные T3SS белки, называемые внешними белками Yersinia (Yops), включают Yop B / D, которые образуют поры в мембране клетки-хозяина и связаны с цитолизом . YopO, YopH , YopM, YopT, YopJ и YopE вводятся в цитоплазму клеток-хозяев с помощью T3SS в пору, частично созданную YopB и YopD. [29] Введенные Yops ограничивают фагоцитоз и клеточные сигнальные пути, важные для врожденной иммунной системы , как обсуждается ниже. Кроме того, некоторые Y. pestisштаммы способны вмешиваться в передачу иммунных сигналов (например, предотвращая высвобождение некоторых цитокинов ).
Y. pestis размножается внутри лимфатических узлов , где он может избежать разрушения клетками иммунной системы, такими как макрофаги . Способность Y. pestis подавлять фагоцитоз позволяет ему расти в лимфатических узлах и вызывать лимфаденопатию . YopH - это протеинтирозинфосфатаза, которая способствует способности Y. pestis уклоняться от клеток иммунной системы. [30] В макрофагах YopH дефосфорилирует p130Cas , Fyb ( Fyn- связывающий белок), SKAP-HOM и Pyk , тирозинкиназу.гомологичен FAK . YopH также связывает субъединицу p85 фосфоинозитид-3-киназы , Gab1 , адаптерные белки Gab2 и фактор обмена гуаниновых нуклеотидов Vav .
YopE функционирует как белок, активирующий GTPase для членов семейства Rho GTPases, таких как RAC1 . YopT - это цистеиновая протеаза, которая ингибирует RhoA , удаляя изопренильную группу , которая важна для локализации белка на клеточной мембране . Было высказано предположение, что YopE и YopT действуют для ограничения YopB / D-индуцированного цитолиза. [31] Это может ограничивать функцию YopB / D по созданию пор, используемых для встраивания Yop в клетки-хозяева, и предотвращать YopB / D-индуцированный разрыв клеток-хозяев и высвобождение содержимого клетки, которое могло бы привлекать и стимулировать ответы иммунной системы.
YopJ является ацетилтрансферазами , который связывается с консервативным альфа-спиралью из МАРК киназ . [32] YopJ ацетилирует киназы MAPK по серинам и треонинам , которые обычно фосфорилируются во время активации каскада киназ MAP . [33] [34] YopJ активируется в эукариотических клетках при взаимодействии с фитиновой кислотой клетки-мишени (IP6). [35] Это нарушение активности протеинкиназы клетки-хозяина вызывает апоптоз.макрофагов, и предполагается, что это важно для установления инфекции и уклонения от иммунного ответа хозяина. YopO представляет собой протеинкиназу, также известную как протеинкиназа A Yersinia (YpkA). YopO является мощным индуктором апоптоза макрофагов человека. [36]
Также было высказано предположение, что бактериофаг - Ypφ - мог быть ответственным за повышение вирулентности этого организма. [37]
В зависимости от того, какой формой чумы заражается человек, чума переходит в другое заболевание; однако чума в целом влияет на способность клетки-хозяина взаимодействовать с иммунной системой, не позволяя организму доставлять фагоцитарные клетки в область инфекции.
Y. pestis - универсальный убийца. Известно, что помимо грызунов и людей он убивал верблюдов, кур и свиней. [38] Домашние собаки и кошки также восприимчивы к чуме, но кошки с большей вероятностью заболеют при заражении. В любом случае симптомы аналогичны тем, которые испытывают люди, и могут быть смертельными для животного. Люди могут подвергнуться воздействию при контакте с инфицированным животным (живым или мертвым) или при вдыхании инфекционных капель, которые больная собака или кошка кашлянула в воздух. [39] [40]
Иммунитет [ править ]
Формалина -inactivated вакцины в прошлом [ когда? ], доступный в Соединенных Штатах для взрослых с высоким риском заражения чумой, пока он не будет снят с продажи Управлением по контролю за продуктами и лекарствами . Он имел ограниченную эффективность и мог вызвать сильное воспаление . Эксперименты с генной инженерией вакцины на основе антигенов F1 и V продолжаются и показывают многообещающие результаты. Однако бактерии, лишенные антигена F1, все еще вирулентны, а антигены V достаточно вариабельны, так что вакцины, состоящие из этих антигенов, могут не обладать полной защитой. [41]Медицинский научно-исследовательский институт инфекционных заболеваний армии США обнаружил, что экспериментальная вакцина на основе антигена F1 / V защищает крабоядных макак , но не защищает африканские зеленые обезьяны . [42] Систематический обзор Кокрановского сотрудничества не выявил исследований достаточного качества, чтобы сделать какое-либо заявление об эффективности вакцины. [43]
Изоляция и идентификация [ править ]
В 1894 году два бактериолога, Александр Йерсин из Швейцарии и Китасато Шибасабуро из Японии, независимо изолировали в Гонконге бактерию, вызвавшую гонконгскую чуму 1894 года . Хотя оба исследователя сообщили о своих выводах, серия сбивающих с толку и противоречивых заявлений Китасато в конечном итоге привела к тому, что Йерсин был признан главным исследователем этого организма. Йерсен назвал его Pasteurella pestis в честь института Пастера , где он работал. В 1967 году переведен в новый род и переименован в Yersinia pestis.в его честь. Йерсин также отметил, что крысы поражались чумой не только во время эпидемий чумы, но также часто предшествовали таким эпидемиям у людей, и что чума рассматривалась многими местными жителями как болезнь крыс; жители деревень в Китае и Индии утверждали, что, когда было обнаружено большое количество мертвых крыс, вскоре последовали вспышки чумы. [ необходима цитата ]
В 1898 году французский ученый Поль-Луи Симон (который также приехал в Китай для борьбы с Третьей пандемией) обнаружил переносчика крысы и блох, который вызывает болезнь. Он отметил, что заболевшим людям необязательно находиться в тесном контакте друг с другом, чтобы заразиться. В Юньнани , Китай, жители покидали свои дома, как только видели дохлых крыс, а на острове Формоза ( Тайвань)), жители считали, что обращение с мертвыми крысами повышает риск развития чумы. Эти наблюдения заставили его подозревать, что блохи могут быть промежуточным фактором в передаче чумы, поскольку люди заражаются чумой только в том случае, если они контактировали с крысами, умершими менее чем за 24 часа до этого. В ставшем уже классическим эксперименте Саймонд продемонстрировал, как здоровая крыса умерла от чумы после того, как зараженные блохи прыгнули на нее от крысы, недавно умершей от чумы. [44] Вспышка распространилась на Чайнатаун, Сан-Франциско, с 1900 по 1904 год, а затем на Окленд и Восточный залив с 1907 по 1909 год. [45]С тех пор он присутствует у грызунов в западной части Северной Америки, поскольку из-за страха перед последствиями вспышки для торговли власти вынуждены прятать мертвых жителей китайского квартала достаточно долго, чтобы болезнь передалась широко распространенным видам местных грызунов в отдаленных районах. области. [46]
Доказательства древней ДНК [ править ]
В 2018 году было опубликовано появление и распространение патогена во время упадка неолита (еще 6000 лет назад). [2] Место в Швеции было источником доказательств ДНК, и торговые сети были предложены как вероятный путь распространения, а не миграции популяций.
Доказательства ДНК, опубликованные в 2015 году, указывают на то, что Y. pestis заразил людей 5000 лет назад в Евразии бронзового века [47], но генетические изменения, которые сделали его очень вирулентным, произошли только около 4000 лет назад. [48] Высоковирулентная версия, способная передаваться блохами через грызунов, людей и других млекопитающих, была обнаружена у двух лиц, связанных с культурой Срубная, из Самарской области в России примерно 3800 лет назад и у человека железного века из Капана , Армения. примерно 2900 лет назад. [48] [47] Это указывает на то, что по крайней мере две линии Y. pestisциркулировали в эпоху бронзы в Евразии. [48] Ю. Pestis бактерия имеет относительно большое число нефункционирующих генов и трех «неуклюжие» плазмид, что указывает на происхождение менее чем 20000 лет назад. [38]
Различают три основных штамма: Y. p. antiqua , вызвавшая пандемию чумы в шестом веке; Ю. П. средневековье , вызвавшее Черную смерть и последующие эпидемии во время второй волны пандемии; и Ю. orientalis , ответственный за текущие вспышки чумы. [49]
Последние события [ править ]
В 2008 году чума обычно обнаруживалась в странах Африки к югу от Сахары и на Мадагаскаре, в районах, на которые приходилось более 95% зарегистрированных случаев. [5]
В сентябре 2009 года смерть Малькольма Касадабана , профессора молекулярной генетики Чикагского университета , была связана с его работой над ослабленным лабораторным штаммом Y. pestis . [50] Было высказано предположение, что гемохроматоз является предрасполагающим фактором смерти Касадабана от этого ослабленного штамма, использованного в исследованиях. [51]
В 2010 году исследователи из Германии определенно установили, используя доказательства ПЦР из образцов, полученных от жертв Черной смерти, что Y. pestis была причиной средневековой Черной смерти . [52]
В 2011 году был опубликован первый геном Y. pestis, выделенный от жертв черной смерти, и сделан вывод о том, что этот средневековый штамм является предком большинства современных форм Y. pestis . [53]
В 2015 году Cell опубликовал результаты исследования древних могил. [54] Плазмиды из Y. Pestis были обнаружены в археологических образцах зубов семи бронзы лиц, в Афанасьево культуры в Сибири, культуры шнуровой керамики в Эстонии, культуры синташтинской в России, культуре Únětice в Польше, и Андроновская культура в Сибири. [55]
8 сентября 2016 года бактерия Y. pestis была идентифицирована по ДНК в зубах, найденных на строительной площадке Crossrail в Лондоне . Человеческие останки оказались жертвами Великой лондонской чумы , которая длилась с 1665 по 1666 год [56].
15 января 2018 года исследователи из Университета Осло и Университета Феррары предположили, что люди и их паразиты были крупнейшими переносчиками чумы. [57] [58]
3 ноября 2019 года в больнице в районе Чаоян в Пекине были диагностированы два случая легочной чумы , что вызвало опасения относительно вспышки. Врачи диагностировали лихорадку у мужчины средних лет, который жаловался на затрудненное дыхание в течение примерно десяти дней в сопровождении своей жены с аналогичными симптомами. [59] Полиция поместила в карантин отделение неотложной помощи в больнице, а китайские агрегаторы новостей установили контроль. [59] 18-го числа был зарегистрирован третий случай заболевания у 55-летнего мужчины из Ксилингольской лиги , одного из двенадцати монгольских автономных регионов в Северном Китае. Пациент получил лечение, и 28 человек без симптомов были помещены в карантин. [60]
В июле 2020 года официальные лица ужесточили меры предосторожности после того, как в Баяннуре , городе в автономном районе Внутренняя Монголия Китая, был подтвержден случай бубонной чумы . Больной был помещен на карантин и пролечен. По данным China's Global Times , второй подозреваемый случай также был расследован, и было объявлено предупреждение 3-го уровня, действующее до конца года. Он запретил охоту и употребление в пищу животных, которые могут быть переносчиками чумы, и призвал общественность сообщать о подозреваемых случаях заболевания. [61]
Ссылки [ править ]
- ^ Райан KJ, Рэй CG, ред. (2004). Шеррис Медицинская микробиология (4-е изд.). Макгроу Хилл. стр. 484 -488. ISBN 978-0-8385-8529-0.
- ^ a b Раскован, Николас; Сьёгрен, Карл-Йоран; Кристиансен, Кристиан; Нильсен, Расмус; Виллерслев, Эске; Desnues, Кристель; Расмуссен, Саймон (2019). «Возникновение и распространение базальных линий Yersinia pestis во время упадка неолита» . Cell . 176 (1–2): 295–305.e10. DOI : 10.1016 / j.cell.2018.11.005 . PMID 30528431 .
- ^ Bockemühl J (1994). «100 лет спустя после открытия возбудителя чумы - важность и почитание Александра Йерсена во Вьетнаме сегодня». Immun Infekt . 22 (# 2): 72–75. PMID 7959865 .
- Перейти ↑ Howard-Jones N (1973). «Был ли Китасато Шибасабуро первооткрывателем бациллы чумы?». Perspect Biol Med . 16 (# 2): 292–307. DOI : 10,1353 / pbm.1973.0034 . PMID 4570035 . S2CID 31767623 .
- ^ a b CDC, «Чума» , Центры по контролю и профилактике заболеваний, октябрь 2017 г.
 Эта статья включает текст из этого источника, который находится в общественном достоянии .
Эта статья включает текст из этого источника, который находится в общественном достоянии . - ^ а б Коллинз FM (1996). Барон С; и другие. (ред.).Пастерелла , Иерсиния и Франсиселла . В: Медицинская микробиология Барона (4-е изд.). Univ. Техасского медицинского отделения. ISBN 978-0-9631172-1-2.
- ^ Stackebrandt, Erko; Дворкин, Мартин; Фалькоу, Стэнли; Розенберг, Евгений; Карл-Хайнц Шлейфер (2005). Прокариоты: Справочник по биологии бактерий: Том 6: Протеобактерии: подкласс гамма . Берлин: Springer. ISBN 978-0-387-25499-9.
- ^ Дэн, Вт .; Burland, V .; Plunkett III, G .; Boutin, A .; Mayhew, GF; Liss, P .; Perna, NT; Роза, диджей; Mau, B .; Чжоу, S .; Шварц, округ Колумбия; Fetherston, JD; Lindler, LE; Брубейкер, Р.Р .; Плано, Г.В. Стрейли, Южная Каролина; McDonough, KA; Nilles, ML; Matson, JS; Блаттнер, Франция; Перри, Р. Д. (август 2002 г.). «Последовательность генома Yersinia pestis KIM» . Журнал бактериологии . 184 (# 16): 4601–4611. DOI : 10.1128 / JB.184.16.4601-4611.2002 . PMC 135232 . PMID 12142430 .
- ^ Parkhill, J .; Wren, BW; Томсон, штат Северная Каролина; Титболл, RW; Холден, HT; Прентис, МБ; Себайхия, М .; Джеймс, KD; Churcher, C .; Мунгалл, КЛ; Baker, S .; Basham, D .; Бентли, SD; Brooks, K .; Cerdeño-Tárraga, AM; Chillingworth, T .; Cronin, A .; Дэвис, РМ; Davis, P .; Dougan, G .; Feltwell, T .; Hamlin, N .; Holroyd, S .; Jagels, K .; Карлышев А.В.; Кожа, S .; Moule, S .; Ойстон, ПК; Перепел, М .; Rutherford, K .; Simmonds, M .; Skelton, J .; Стивенс, К .; Whitehead, S .; Баррелл, Б.Г. (октябрь 2001 г.). «Последовательность генома Yersinia pestis , возбудителя чумы» . Природа . 413 (# 6, 855): 523–527. Bibcode : 2001Natur.413..523P . doi :10.1038 / 35097083 . PMID 11586360 .
- ^ Цепь PS, Ху П., Малфатти С.А. и др. (2006). «Полная последовательность генома штаммов Yersinia pestis Antiqua и Nepal 516: доказательства уменьшения гена у появляющегося патогена» . J. Bacteriol . 188 (# 12): 4453–4463. DOI : 10.1128 / JB.00124-06 . PMC 1482938 . PMID 16740952 .
- ^ a b Hinnebusch BJ, Rudolph AE, Cherepanov P, Dixon JE, Schwan TG, Forsberg A (2002). «Роль мышиного токсина Yersinia в выживании Yersinia pestis в средней кишке переносчика блох». Наука . 296 (# 5, 568): 733–735. Bibcode : 2002Sci ... 296..733H . DOI : 10.1126 / science.1069972 . PMID 11976454 . S2CID 34770234 .
- ^ a b Lathem WW, Price PA, Miller VL, Goldman WE (2007). «Активирующая плазминоген протеаза специфически контролирует развитие первичной легочной чумы». Наука . 315 (# 5, 811): 509–513. Bibcode : 2007Sci ... 315..509L . DOI : 10.1126 / science.1137195 . PMID 17255510 . S2CID 39881239 .
- ^ Хиксон, KK; Адкинс, JN; Baker, SE; Мур, Р.Дж.; Хроми, BA; Смит, Р. Д.; Маккатчен-Мэлони, SL; Липтон, MS (ноябрь 2006 г.). «Идентификация кандидата в биомаркеры в Yersinia pestis с использованием полуколичественной протеомики в масштабах всего организма». Журнал протеомных исследований . 5 (# 11): 3008–3017. DOI : 10.1021 / pr060179y . PMID 17081052 .
- ^ Скиано, Челси А .; Ку, Йованка Т .; Schipma, Matthew J .; Колфилд, Адам Дж .; Джафари, Надере; Латем, Уиндем У. (01.05.2014). «Полногеномный анализ малых РНК, экспрессируемых Yersinia pestis, идентифицирует регулятор системы секреции Yop-Ysc типа III» . Журнал бактериологии . 196 (# 9): 1659–1670. DOI : 10.1128 / JB.01456-13 . ISSN 1098-5530 . PMC 3993326 . PMID 24532772 .
- ^ Корнелис, GR; Boland, A .; Бойд, AP; Geuijen, C .; Ириарте, М .; Neyt, C .; Сори, депутат; Stainier, I. (1998-12-01). «Плазмида вирулентности Yersinia, антихозяинный геном» . Обзоры микробиологии и молекулярной биологии . 62 (# 4): 1315–1352. DOI : 10.1128 / MMBR.62.4.1315-1352.1998 . ISSN 1092-2172 . PMC 98948 . PMID 9841674 .
- ^ Ян, Яньфэн; Су, Шаньчунь; Мэн, Сянжун; Цзи, Сяолань; Цюй, Йи; Лю, Цзычжун; Ван, Сяои; Цуй, Юйцзюнь; Дэн, Чжунлян (2013). «Определение экспрессии sRNA с помощью RNA-seq в Yersinia pestis, выращенной in vitro и во время инфекции» . PLOS ONE . 8 (# 9): e74495. Bibcode : 2013PLoSO ... 874495Y . DOI : 10.1371 / journal.pone.0074495 . ISSN 1932-6203 . PMC 3770706 . PMID 24040259 .
- ^ Мейер KF (1957). «Естественная история чумы и орнитоза: лекция Р. Э. Дайера» . Отчеты об общественном здравоохранении . 72 (# 8): 705–719. DOI : 10.2307 / 4589874 . JSTOR 4589874 . PMC 2031327 . PMID 13453634 .
- ^ фон Рейн С.Ф., Вебер Н.С., Темпест Б. и др. (1977). «Эпидемиологические и клинические особенности вспышки бубонной чумы в Нью-Мексико». J. Infect. Дис . 136 (# 4): 489–494. DOI : 10.1093 / infdis / 136.4.489 . PMID 908848 .
- ^ Pauli JN, Buskirk SW, Williams ES, Edwards WH (2006). «Эпизоотия чумы у чернохвостой луговой собачки (Cynomys ludovicianus)». J. Wildl. Дис . 42 (# 1): 74–80. DOI : 10.7589 / 0090-3558-42.1.74 . PMID 16699150 . S2CID 9716200 .
- Перейти ↑ Webb CT, Brooks CP, Gage KL, Antolin MF (2006). «Классическая передача блох не вызывает эпизоотий чумы у луговых собачек» . Proc. Natl. Акад. Sci. США . 103 (# 16): 6236–6241. Bibcode : 2006PNAS..103.6236W . DOI : 10.1073 / pnas.0510090103 . PMC 1434514 . PMID 16603630 .
- ^ Bertherat, E .; Бехуча, С .; Chougrani, S .; Разик, Ф .; Duchemin, JB; Houti, L .; Deharib, L .; Fayolle, C .; Makrerougrass, B .; Дали-Яхья, Р .; Bellal, R .; Belhabri, L .; Chaieb, A .; Тихомиров, Э .; Карниэль, Э. (октябрь 2007 г.). «Повторное появление чумы в Алжире через 50 лет, 2003 г.» . Возникающие инфекционные заболевания . 13 (# 10): 1459–1462. DOI : 10.3201 / eid1310.070284 . PMC 2851531 . PMID 18257987 .
- Перейти ↑ Zhou D, Han Y, Yang R (2006). «Молекулярное и физиологическое понимание передачи, вирулентности и этиологии чумы». Микробы заражают . 8 (# 1): 273–284. DOI : 10.1016 / j.micinf.2005.06.006 . PMID 16182593 .
- ^ Hinnebusch, BJ; Перри, Р. Д. и Шван, Т. Г. (1996). «Роль локуса накопления гемина Yersinia pestis (hms) в передаче чумы блохами». Наука . 273 (# 5, 237): 367–370. Bibcode : 1996Sci ... 273..367H . DOI : 10.1126 / science.273.5273.367 . PMID 8662526 . S2CID 37512575 .
- ^ Эриксон, DL; NR Waterfield; В. Вадивалоо; Д. Лонг; Э. Р. Фишер; Р. Френч-Констан и Б. Дж. Хиннебуш (2007). «Острая оральная токсичность Yersinia pseudotuberculosis для блох: последствия для эволюции трансмиссивной передачи чумы» . Клеточная микробиология . 9 (# 11): 2658–2666. DOI : 10.1111 / j.1462-5822.2007.00986.x . PMID 17587333 . S2CID 36769530 .
- ^ Джарретт, Колорадо; Deak, E; Isherwood, KE; Ойстон, ПК; Фишер, ER; Уитни, Арканзас; Кобаяши, SD; ДеЛео, Франция; Хиннебуш, Б. Дж. (2004). «Передача Yersinia pestis из инфекционной биопленки в переносчике блох» . J Infect Dis . 190 (# 4): 783–792. DOI : 10.1086 / 422695 . PMID 15272407 .
- ^ Эриксон DL, Джарретт Колорадо; Wren, BW; Хиннебуш, Б. Дж. (2006). «Различия серотипов и отсутствие образования биопленок характеризуют инфекцию Yersinia pseudotuberculosis, вызванную блохами- переносчиками Xenopsylla cheopis Yersinia pestis » . J Bacteriol . 188 (# 3): 1113–1119. DOI : 10.1128 / jb.188.3.1113-1119.2006 . PMC 1347331 . PMID 16428415 .
- ^ а б Hinnebusch, BJ; Эриксон, DL (2008). «Биопленка Yersinia pestis в переносчике блох и ее роль в передаче чумы» . Актуальные темы микробиологии и иммунологии . 322 : 229–248. DOI : 10.1007 / 978-3-540-75418-3_11 . ISBN 978-3-540-75417-6. ISSN 0070-217X . PMC 3727414 . PMID 18453279 .
- ^ Salyers AA, Витт DD (2002). Бактериальный патогенез: молекулярный подход (2-е изд.). ASM Press. С. 207–212.
- ^ Viboud GI, Bliska JB (2005). «Внешние белки иерсинии: роль в модуляции сигнальных ответов клетки-хозяина и патогенезе». Анну. Rev. Microbiol . 59 (# 1): 69–89. DOI : 10.1146 / annurev.micro.59.030804.121320 . PMID 15847602 .
- ↑ de la Puerta ML, Trinidad AG, del Carmen Rodríguez M, Bogetz J, Sánchez Crespo M, Mustelin T, Alonso A, Bayón Y (февраль 2009 г.). Бозза П. (ред.). «Характеристика новых субстратов, нацеленных на тирозинфосфатазу Yersinia YopH» . PLOS ONE . 4 (# 2): e4431. Bibcode : 2009PLoSO ... 4.4431D . DOI : 10.1371 / journal.pone.0004431 . PMC 2637541 . PMID 19221593 .
- ^ Мехия E, Bliska JB, Viboud GI (февраль 2009). « Yersinia контролирует доставку эффектора типа III в клетки-хозяева путем модуляции активности Rho» . PLOS ONE . 4 (# 2): e4431. DOI : 10.1371 / journal.ppat.0040003 . PMC 2186360 . PMID 18193942 .
- ↑ Hao YH, Wang Y, Burdette D, Mukherjee S, Keitany G, Goldsmith E, Orth K (январь 2008 г.). Коби Б. (ред.). «Структурные требования для Yersinia YopJ ингибирования путей MAP киназы» . PLOS ONE . 3 (# 1): e1375. Bibcode : 2008PLoSO ... 3.1375H . DOI : 10.1371 / journal.pone.0001375 . PMC 2147050 . PMID 18167536 .
- ^ Mukherjee, S .; Кейтани, Глэдис; Ли, Ян; Ван, Юн; Болл, Гайдн Л .; Голдсмит, Элизабет Дж .; Орт, Ким (2006). « Yersinia YopJ Acetylates и ингибирует активацию киназы путем блокирования фосфорилирования». Наука . 312 (# 5, 777): 1211–1214. Bibcode : 2006Sci ... 312.1211M . DOI : 10.1126 / science.1126867 . PMID 16728640 . S2CID 13101320 .
- ^ Mittal, R .; Peak-Chew, S.-Y .; МакМахон, HT (2006). «Ацетилирование остатков петли активации MEK2 и IB-киназы (IKK) с помощью YopJ ингибирует передачу сигналов» . Труды Национальной академии наук . 103 (# 49): 18574–18579. Bibcode : 2006PNAS..10318574M . DOI : 10.1073 / pnas.0608995103 . PMC 1654131 . PMID 17116858 .
- Перейти ↑ Mittal R, Peak-Chew SY, Sade RS, Vallis Y, McMahon HT (2010). «Ацетилтрансферазная активность бактериального токсина YopJ Yersinia активируется эукариотической клеткой-хозяином инозитом гексакисфосфатом» . J Biol Chem . 285 (№ 26): 19927–19934. DOI : 10.1074 / jbc.M110.126581 . PMC 2888404 . PMID 20430892 .
- ^ Парк H, Тея K, О'Ши JJ, Siegel RM (май 2007). « Эффекторный белок Yersinia YpkA индуцирует апоптоз независимо от деполимеризации актина» . J. Immunol . 178 (# 10): 6426–6434. DOI : 10.4049 / jimmunol.178.10.6426 . PMID 17475872 .
- ^ Дербис, А; Chenal-Francisque, V; Pouillot, F; Fayolle, C; Прево, MC; Médigue, C; Hinnebusch, BJ; Карниэль, Э (2007). «Приобретенный горизонтально нитчатый фаг вносит свой вклад в патогенность бациллы чумы» . Mol Microbiol . 63 (# 4): 1145–1157. DOI : 10.1111 / j.1365-2958.2006.05570.x . PMID 17238929 . S2CID 30862265 .
- ^ a b Келли, Джон (2005). Великая смертность: интимная история Черной смерти (1-е изд.). Лондон [ua]: Четвертое сословие. п. 35. ISBN 978-0007150694.
- ^ «Кошки - здоровые животные, здоровые люди» . Центры по контролю и профилактике заболеваний. 2016-05-13 . Проверено 25 ноября 2016 .
- ^ «Собаки - здоровые животные, здоровые люди» . Центры по контролю и профилактике заболеваний. 2020-02-21 . Проверено 30 июня 2020 .
- ^ Welkos S, et al. (2002). «Определение вирулентности штаммов Yersinia pestis с дефицитом пигментации и активатора пигментации / плазминогена на моделях легочной чумы приматов и мышей, не относящихся к человеку». Вакцина . 20 (17–18): 2206–2214. DOI : 10.1016 / S0264-410X (02) 00119-6 . PMID 12009274 .
- ↑ Pitt ML (13 октября 2004 г.). Нечеловеческие приматы как модель легочной чумы (PDF) . Семинар по моделям животных и коррелятам защиты от чумных вакцин, Гейтерсбург, Мэриленд. Центр оценки и исследований биологических препаратов (Управление по контролю за продуктами и лекарствами, Департамент здравоохранения и человеческих ресурсов). С. 222–248. Архивировано из оригинального (PDF) 25 декабря 2004 года.
- ^ Джефферсон Т, Demicheli В, Пратт М (2000). Джефферсон Т. (ред.). «Вакцины для профилактики чумы» . Кокрановская база данных Syst Rev (2): CD000976. DOI : 10.1002 / 14651858.CD000976 . ISSN 1465-1858 . PMC 6532692 . PMID 10796565 . Изобразительное искусство. № CD000976.
- ^ "Чума" . Ассоциация Amicale Sante Navale et d'Outremer. Архивировано из оригинала на 4 сентября 2012 года.
- ^ "В этот день: Начало вспышки бубонной чумы в Сан-Франциско" . В поисках Дульсинеи . Проверено 25 ноября 2017 .
- Перейти ↑ Chase, M. (2004). Берберийская чума: Черная смерть в викторианском Сан-Франциско . Произвольная торговля в мягкой обложке.
- ^ a b Расмуссен, Саймон; Аллентофт, Мортен Эрик; Нильсен, Каспер; Орландо, Людовик; Сикора, Мартин; Сьёгрен, Карл-Йоран; Педерсен, Андерс Горм; Шуберт, Миккель; Ван Дам, Алекс; Капель, Кристиан Мольин Аутцен; Нильсен, Хенрик Бьёрн; Брунак, Сорен; Аветисян, Павел; Епимахов Андрей; Халяпин Михаил Викторович; Гнуни, Артак; Крийска, Айвар; Ласак, Ирена; Мецпалу, Майт; Моисеев, Вячеслав; Громов, Андрей; Покутта, Далия; Сааг, Лехти; Варул, Лийви; Епископосян, Левон; Зихериц-Понтен, Томас; Фоли, Роберт А.; Лар, Марта Мирасон; Нильсен, Расмус; Кристиансен, Кристиан; Виллерслев, Эске (2015). "Ранние расходящиеся штаммы Yersinia pestis в Евразии 5000 лет назад" . Cell . 163 (# 3): 571–582. Дои: 10.1016 / j.cell.2015.10.009 . ISSN 0092-8674 . PMC 4644222 . PMID 26496604 . Эта статья содержит цитаты из этого источника, доступного по лицензии Creative Commons Attribution 4.0 International (CC BY 4.0) .
- ^ a b c Спайроу, Мария А .; Тухбатова, Резеда И .; Ван, Чуан-Чао; а; Lankapalli, Aditya K .; Кондрашин, Виталий В .; Цыбин Виктор А .; Хохлов Александр; hnert; Хербиг, Александр; Бос, Кирстен I .; Краузе, Йоханнес (2018-06-08). «Анализ геномов Yersinia pestis возрастом 3800 лет позволяет предположить происхождение бубонной чумы бронзового века» . Nature Communications . 9 (# 1): 2234. Bibcode : 2018NatCo ... 9.2234S . DOI : 10.1038 / s41467-018-04550-9 . ISSN 2041-1723 . PMC 5993720 . PMID 29884871 . Эта статья содержит цитаты из этого источника, доступного по лицензии Creative Commons Attribution 4.0 International (CC BY 4.0) .
- ^ Ахтман, М; Zurth, K; Морелли, G; Торреа, G; Guiyoule, A; Карниэль, Э (1999). « Yersinia pestis , вызывающая чуму, представляет собой недавно появившийся клон Yersinia pseudotuberculosis» . Proc Natl Acad Sci USA . 96 (# 24): 14043–14048. Bibcode : 1999PNAS ... 9614043A . DOI : 10.1073 / pnas.96.24.14043 . PMC 24187 . PMID 10570195 .
- ^ Sadovi, Карлос (2009-09-19). «Исследователь U. of C. умирает от заражения чумными бактериями» . Чикагский центр последних новостей . Проверено 3 марта 2010 .
- ↑ Рэндалл, Том (25 февраля 2011 г.). «Смерть от чумы наступила в течение нескольких часов, спровоцированная состоянием здоровья ученого» .
- ^ Haensch S, Bianucci R, Signoli M, Rajerison M, Schultz M, Kacki S, Vermunt M, Weston DA, Hurst D, Achtman M, Carniel E, Bramanti B (2010). «Определенные клоны Yersinia pestis вызвали черную смерть» . PLOS Патогены . 6 (# 10): e1001134. DOI : 10.1371 / journal.ppat.1001134 . PMC 2951374 . PMID 20949072 .
- ↑ Bos KI , Schuenemann VJ, Golding GB, Burbano HA, Waglechner N, Coombes BK, McPhee JB, DeWitte SN, Meyer M, Schmedes S, Wood J, Earn DJ, Herring DA, Bauer P, Poinar HN, Krause J (12 Октябрь 2011 г.). «Проект генома Yersinia pestis от жертв Черной смерти» . Природа . 478 (7370): 506–10. Bibcode : 2011Natur.478..506B . DOI : 10,1038 / природа10549 . PMC 3690193 . PMID 21993626 .
- ^ Расмуссен, Саймон; Аллентофт, Мортен Эрик; Нильсен, Каспер; Орландо, Людовик; Сикора, Мартин; Сьёгрен, Карл-Йоран; Педерсен, Андерс Горм; Шуберт, Миккель; Ван Дам, Алекс; Капель, Кристиан Мольин Аутцен; Нильсен, Хенрик Бьёрн; Брунак, Сорен; Аветисян, Павел; Епимахов Андрей; Халяпин Михаил Викторович; Гнуни, Артак; Крийска, Айвар; Ласак, Ирена; Мецпалу, Майт; Моисеев, Вячеслав; Громов, Андрей; Покутта, Далия; Сааг, Лехти; Варул, Лийви; Епископосян, Левон; Зихериц-Понтен, Томас; Фоли, Роберт А.; Лар, Марта Мирасон; Нильсен, Расмус; и другие. (2015). "Ранние расходящиеся штаммы Yersinia pestis в Евразии 5000 лет назад" . Cell . 163 (# 3): 571–582. DOI : 10.1016 / j.cell.2015.10.009 .PMC 4644222 . PMID 26496604 .
- Перейти ↑ Zimmer, Carl (2015-10-22). «В древней ДНК доказательства чумы гораздо раньше, чем было известно» . Нью-Йорк Таймс . ISSN 0362-4331 . Проверено 16 апреля 2020 .
- ^ «Впервые идентифицирована ДНК бактерий, ответственных за Великую чуму в Лондоне 1665 года» .
- ^ «Не вините крыс: человеческие блохи и вши, вероятно, распространяют черную смерть» . CBC News .
- ^ Дин, Кэтрин R; Крауэр, Фабьен; Валло, Ларс; Лингьерде, Уле Кристиан; Браманти, Барбара; Стенсет, Нильс Хр; Шмид, Борис V (2018). «Человеческие эктопаразиты и распространение чумы в Европе во время Второй пандемии» . Труды Национальной академии наук . 115 (# 6): 1304–1309. DOI : 10.1073 / pnas.1715640115 . PMC 5819418 . PMID 29339508 .
- ^ а б Ви, Суи-Ли (13.11.2019). «В Китае диагностирована легочная чума» . Нью-Йорк Таймс . ISSN 0362-4331 . Проверено 13 ноября 2019 .
- ^ «Бубонная чума: Третий случай зарегистрирован в Китае» .
- ^ «Китайская бубонная чума: Внутренняя Монголия принимает меры предосторожности после каждого случая» . BBC. 2020-07-06 . Проверено 6 июля 2020 .
Внешние ссылки [ править ]
| Викискладе есть медиафайлы по теме Yersinia pestis . |
| В Wikispecies есть информация о Yersinia pestis . |
- Yersinia pestis . Виртуальный музей бактерий.
- Список разновидностей штаммов и информация о синонимах (и многое другое) доступны через браузер таксономии NCBI .
- Домашняя страница CDC, посвященная чуме
- Страница ресурсов IDSA по чуме : актуальная, исчерпывающая информация о патогенезе, микробиологии, эпидемиологии, диагностике и лечении
- Чума (Yersinia Pestis)
- Wyndham Lathem, выступающий на тему «От легкой степени к смертоносной: как Yersinia pestis эволюционировала, чтобы вызвать легочную чуму».