| Диктиостелиды | |
|---|---|
 | |
| Dictyostelium discoideum | |
| Научная классификация | |
| Домен: | Эукариоты |
| Тип: | Амебозоа |
| Подтип: | Коноса |
| Инфрафилюм: | Mycetozoa |
| Учебный класс: | Диктиостелия |
| Заказы | |
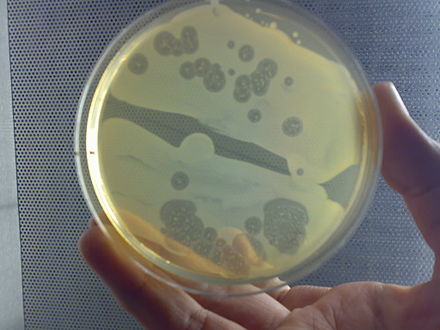
В dictyostelids ( Dictyostelia / Dictyostelea , МКЗН или Dictyosteliomycetes , ICBN ) представляют собой группу клеточных форм слизи или социальной амебы .
Многоклеточное поведение [ править ]
{kind=link}
{kind=link}
Когда пища (обычно бактерии) легкодоступна, диктиостелиды ведут себя как отдельные амебы, которые нормально питаются и делятся. Однако, когда запасы пищи истощены, они собираются в многоклеточную совокупность, называемую псевдоплазмодием, грексом или слизней (не путать с брюхоногим моллюском, называемым слизеньком ). Грекс имеет четко выраженную переднюю и заднюю части, реагирует на световые и температурные градиенты и обладает способностью к миграции. При правильных обстоятельствах грек созревает, образуя сорокарпий (плодовое тело) со стеблем, поддерживающим один или несколько сорусов.(клубки спор). Эти споры представляют собой неактивные клетки, защищенные устойчивыми клеточными стенками, и становятся новыми амебами, когда пища становится доступной.
В Acytostelium , то sorocarp поддерживается стеблом , состоящее из целлюлозы , но и в других dictyostelids стебель состоит из клеток, иногда занимая большую часть первоначальной амебы. За некоторыми исключениями, эти клетки погибают во время формирования стебля, и существует определенное соответствие между частями грекса и частями плодового тела. Агрегация амеб обычно происходит в сходящихся потоках. Амебы передвигаются с помощью филозных псевдопод , и их привлекают химические вещества, производимые другими амебами. У Dictyostelium агрегация сигнализируется цАМФ , но другие используют другие химические вещества. У вида Dictyostelium purpureum, группировка происходит по родству, а не только по близости.
Используется в качестве модельного организма [ править ]
Dictyostelium использовался в качестве модельного организма в молекулярной биологии и генетике и изучается как пример клеточной коммуникации , дифференциации и запрограммированной гибели клеток . Это также интересный пример эволюции сотрудничества и обмана. [1] [2] [3] Большой объем данных исследований, касающихся D. discoideum , доступен в режиме онлайн на DictyBase .
Механизм агрегации у Dictyostelium [ править ]
Механизм агрегации амеб основан на использовании циклического аденозинмонофосфата (цАМФ) в качестве сигнальной молекулы. Одна клетка, основатель колонии, начинает секретировать цАМФ в ответ на стресс. Другие обнаруживают этот сигнал и реагируют двумя способами:
- Амеба движется навстречу сигналу.
- Амеба выделяет больше цАМФ, чтобы усилить сигнал.
Эффект от этого заключается в передаче сигнала через ближайшую популяцию амеб и вызывает движение внутрь к области с наибольшей концентрацией цАМФ.
Внутри отдельной ячейки механизм выглядит следующим образом:
- Рецепция цАМФ на клеточной мембране активирует G-белок
- G-белок стимулирует аденилатциклазу
- цАМФ диффундирует из клетки в среду
- Внутренний цАМФ инактивирует внешний рецептор цАМФ.
- Другой g-белок стимулирует фосфолипазу C
- IP 3 вызывает высвобождение ионов кальция
- Ионы кальция действуют на цитоскелет, вызывая расширение псевдоподий .
Поскольку внутренняя концентрация цАМФ инактивирует рецептор внешнего цАМФ, отдельная клетка демонстрирует колебательное поведение. Такое поведение создает красивые спирали, наблюдаемые в сходящихся колониях, и напоминает реакцию Белоусова-Жаботинского и двумерные циклические клеточные автоматы .
Геном [ править ]
Полный геном Dictyostelium discoideum был опубликован в журнале Nature в 2005 году генетиком Людвигом Эйхингером и его сотрудниками. [4] Гаплоидный геном содержит около 12 500 генов на 6 хромосомах. Для сравнения: диплоидный геном человека насчитывает 20 000-25 000 генов (представленных дважды) на 23 парах хромосом. Высокий уровень нуклеотидов аденозина и тимидина (~ 77%) приводит к тому, что использование кодонов способствует большему количеству аденозинов и тимидинов в третьей позиции. Тандемные повторы тринуклеотидов в изобилии присутствуют в Dictyostelium , которые у человека вызывают нарушения тринуклеотидных повторов .
Половое размножение [ править ]
Половое развитие может происходить, когда амебоидные клетки испытывают нехватку бактериальной пищи и присутствуют темные влажные условия. [5] Как гетероталлические, так и гомоталлические штаммы Dictostelium могут подвергаться спариванию. Гетероталлическое половое развитие наиболее полно изучено у D. discoideum , а гомоталлическое половое развитие наиболее хорошо изучено у D. mucoroides . [6] Гетероталлические спаривания инициируются слиянием гаплоидных клеток (гамет) двух штаммов противоположного типа спаривания. Это контрастирует с гомоталлическими штаммами, которые, по-видимому, выражают оба типа спаривания. [7]
Спаривание инициируется гаметогенезом, который производит маленькие подвижные гаметы, которые сливаются, образуя небольшую двухъядерную клетку . Затем объем двуядерной клетки увеличивается, образуя гигантскую двухъядерную клетку. По мере роста ядра набухают, а затем сливаются, образуя настоящую диплоидную гигантскую зиготную клетку. По мере того как это происходит, амебы подвергались индуцированному цАМФ хемотаксису по отношению к поверхности гигантских клеток. Это образует клеточный агрегат, и в центре агрегата гигантская зиготная клетка поглощает окружающие амебы. За фагоцитозом следует переваривание проглоченных амеб. Затем зигота образует макроцисту, характеризующуюся окружающей внеклеточной целлюлозной оболочкой. После того, как макроциста сформировалась, она обычно остается бездействующей в течение периода, прежде чем может произойти прорастание. [8] Внутри макроцисты диплоидная зигота претерпевает мейоз, за которым следуют последовательные митотические деления. Когда макроциста прорастает, она высвобождает много гаплоидных амебоидных клеток.
Классификация [ править ]
Первым описанным диктиостелидом был Dictyostelium mucoroides в 1869 году Оскар Брефельд. [9]
Впервые обнаруженный в лесу Северной Каролины в 1935 году, Dictyostelium discoideum сначала был классифицирован как «низшие грибы». и в последующие годы в царства Protoctista , Fungi и Tubulomitochondrae . К 1990-м годам большинство ученых приняли нынешнюю классификацию.
Сейчас большинство считает, что амебозоа образуют отдельную кладу на уровне царства, будучи более близкими как к животным, так и к грибам, чем к растениям. [4]
| Кладограмма Dictyostelia [10] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Класс Dictyostelia Lister 1909 г. Оливковое 1970 [11]
- Род? Калоспейра Арно 1949
- Род? Coenonia van Tieghem 1884 non Vandamme et al. 1999 г.
- Род? Synstelium Baldauf, Sheikh & Thulin 2017
- Заказать Actyosteliales Baldauf, Sheikh & Thulin 2017
- Семья Cavenderiaceae Baldauf, Sheikh & Thulin 2017
- Род Cavenderia Baldauf, Sheikh & Thulin 2017
- Семья Acytosteliidae Raper ex Raper & Quinlan 1958
- Род Acytostelium Raper 1956 г.
- Род Heterostelium Baldauf, Sheikh & Thulin 2017
- Род Rostrostelium Baldauf, Sheikh & Thulin 2017
- Семья Cavenderiaceae Baldauf, Sheikh & Thulin 2017
- Заказать Dictyosteliales Olive ex Kirk, Cannon & David 2001
- Род? Coremiostelium Baldauf, Sheikh & Thulin 2017
- Род? Гелиомикопсис Арно 1949
- Род? Pygmomyces Arnaud 1949
- Род †? Myxomitodes Bengtson et al. 2007 г.
- Род? Rhabdocystis Арно 1949
- Семья Dictyosteliidae Rostafinski 1875 ex Cooke 1877
- Dictyostelium Brefeld 1870 г.
- Полисфондилий Брефельд 1884
- Семья Раперостелиевые Baldauf, Sheikh & Thulin 2017
- Род Hagiwaraea Baldauf, Sheikh & Thulin 2017
- Род Raperostelium Baldauf, Sheikh & Thulin 2017
- Род Speleostelium Baldauf, Sheikh & Thulin 2017
- Род Tieghemostelium Baldauf, Sheikh & Thulin 2017
Модель организма-хозяина Legionella [ править ]
Dictyostelium имеет много общих молекулярных особенностей с макрофагами , человеческим хозяином Legionella . Состав цитоскелета D. discoideum аналогичен составу клеток млекопитающих, так как это процессы, управляемые этими компонентами, такие как фагоцитоз, мембранный перенос, эндоцитарный транзит и сортировка везикул. Как и лейкоциты, D. discoideum обладает хемотаксической способностью. Следовательно, D. discoideum представляет собой подходящую модельную систему для определения влияния различных факторов клетки-хозяина во время инфекций Legionella . [12]
Ссылки [ править ]
- ^ Strassman JE, Zhu Y, и Queller DC. (2000) Альтруизм и социальное мошенничество в социальной амебе Dictyostelium discoideum . Природа
- ^ Dao Д.Н., Кессин RH, и Эннис HL (2000). Обман развития и эволюционная биология Dictyostelium и Myxococcus . Микробиология
- ^ Brännsröm Å и Дикман U (2005). Эволюционная динамика альтруизма и мошенничества среди социальных амеб. Труды Лондонского королевского общества Б.
- ^ a b Eichinger, L .; Pachebat, JA; Glöckner, G .; Раджандрим, Массачусетс; Sucgang, R .; Berriman, M .; Song, J .; Olsen, R .; Szafranski, K .; Xu, Q .; и другие. (2005). «Геном социальной амебы Dictyostelium discoideum» . Природа . 435 (7038): 43–57. DOI : 10,1038 / природа03481 . PMC 1352341 . PMID 15875012 .
- ^ Цветы JM, Ли С. И., Stathos A, G Saxer, Ostrowski Е.А., Queller DC, Штрассман JE, Purugganan MD (июль 2010). «Вариация, пол и социальное сотрудничество: молекулярная популяционная генетика социальной амебы Dictyostelium discoideum» . PLOS Genet . 6 (7): e1001013. DOI : 10.1371 / journal.pgen.1001013 . PMC 2895654 . PMID 20617172 .
- ^ O'Day DH, Keszei A (май 2012). «Сигнализация и секс у социальных амебов». Биол Рев Камб Филос Соц . 87 (2): 313–29. DOI : 10.1111 / j.1469-185X.2011.00200.x . PMID 21929567 .
- Перейти ↑ Robson GE, Williams KL (апрель 1980 г.). «Система спаривания клеточной слизистой плесени Dictyostelium discoideum». Curr. Genet . 1 (3): 229–32. DOI : 10.1007 / BF00390948 . PMID 24189663 .
- ^ Никерсон А. В., Рэпер КБ. Макроцисты в жизненном цикле Dictyostelliaceae II. Прорастание макрокист. Являюсь. J. Bot. 1973 60 (3): 247-254.
- ^ Brefeld, О (1869 г.). "Ein neuer Organismus und der Verwandschaft der Myxomyceten". Abh Seckenberg Naturforsch Ges . 7 : 85–107.
- ^ Шейх, Санея; Тулин, Матс; Кавендер, Джеймс С; Эскаланте, Рикардо; Каваками, Син-ичи; Ладо, Карлос; Ландольт, Джон; Нанджундиа, Видьянанд; Квеллер, Дэвид; Штрассманн, Жанна; Spiegel, Frederick W .; Стивенсон, Стив; Vadell, Eduardo M; Балдауф, Сандра (24 ноября 2017 г.). «Новая классификация диктиостелид» . Протисты . 169 : 1–28. DOI : 10.1016 / j.protis.2017.11.001 .
- ^ Виджаявардене, Налин; Хайд, Кевин; Аль-Ани, LKT; Долатабади, S; Стадлер, Марк; Haelewaters, Дэнни; и другие. (2020). «Очерк грибов и грибовидных таксонов» . Микосфера . 11 : 1060–1456. DOI : 10.5943 / mycosphere / 11/1/8 .
- ^ Брун; и другие. (2008). « Dictyostelium , податливый модельный организм-хозяин для Legionella » .Легионелла : молекулярная микробиология . Caister Academic Press. ISBN 978-1-904455-26-4.
Внешние ссылки [ править ]
| Викискладе есть медиафайлы по теме Dictyosteliida . |
- Диктиостелиум (2007)
- Низкое общество (2004)
- Интернет-ресурс по информатике dictyBase для Dictyostelium
- dictyBase wiki официальный вики-сайт dictyBase
- Проект генома Dictyostelium discoideum
- Dictyostelium discoideum описание, жизненный цикл
| vтеАмебозоа | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Дискоса |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tubulinea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Evosea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||