{kind=link}
В клеточной биологии , фокальные адгезии (также клетка-матрица спайка или FAS ) являются крупными макромолекулами узлов , через которые механическую сила и регуляторные сигналы , передаваемые между внеклеточным матриксом (ECM) и взаимодействующей клеткой. Точнее, фокальные адгезии - это субклеточные структуры, которые опосредуют регуляторные эффекты (т.е. сигнальные события) клетки в ответ на адгезию ECM. [1]
Фокальные адгезии служат механическими связями с ЕСМ и биохимическим сигнальным центром для концентрации и направления многочисленных сигнальных белков в сайтах связывания интегрина и кластеризации.
Структура и функции [ править ]
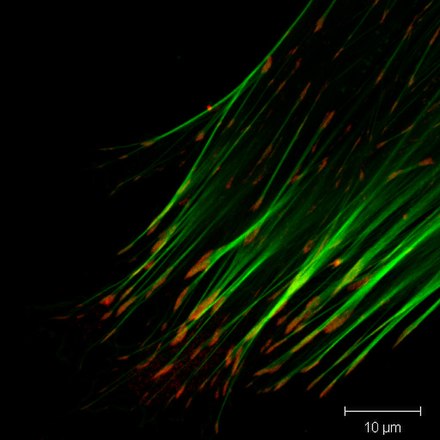
Фокальные адгезии представляют собой содержащие интегрин, мультибелковые структуры, которые образуют механические связи между внутриклеточными пучками актина и внеклеточным субстратом во многих типах клеток. Фокальные адгезии - это большие динамические белковые комплексы, с помощью которых цитоскелет клетки соединяется с ECM. Они ограничены четко определенными диапазонами клетки, в которых плазматическая мембрана закрывается в пределах 15 нм от субстрата ЕСМ. [2] Очаговые адгезии находятся в состоянии постоянного потока: белки постоянно связываются и отделяются от них, поскольку сигналы передаются в другие части клетки, относящиеся ко всему, от подвижности клетки до клеточного цикла. Фокальные адгезии могут содержать более 100 различных белков, что предполагает значительное функциональное разнообразие. [3] Они не только закрепляют ячейку, но и действуют как носители сигнала (датчики), которые информируют ячейку о состоянии ECM и, таким образом, влияют на их поведение. [4] В сидячих клетках фокальные адгезии довольно стабильны в нормальных условиях, в то время как в движущихся клетках их стабильность снижена: это связано с тем, что в подвижных клетках фокальные адгезии постоянно собираются и разбираются, поскольку клетка устанавливает новые контакты на переднем крае. , и ломает старые контакты на заднем крае ячейки. Одним из примеров их важной роли является иммунная система , в которой белые кровяные тельцамигрируют по соединительному эндотелию, следуя клеточным сигналам к поврежденной биологической ткани .
Морфология [ править ]
Связь между фокальными адгезиями и белками внеклеточного матрикса обычно включает интегрины . Интегрины связываются с внеклеточными белками через короткие аминокислотные последовательности, такие как мотив RGD (обнаруженный в таких белках, как фибронектин , ламинин или витронектин ) или мотивы DGEA и GFOGER, обнаруженные в коллагене . Интегрины - гетеродимерыкоторые образованы из одной бета и одной альфа-субъединицы. Эти субъединицы присутствуют в разных формах, соответствующие им лиганды классифицируют эти рецепторы на четыре группы: рецепторы RGD, рецепторы ламинина, рецепторы, специфичные для лейкоцитов, и рецепторы коллагена. Внутри клетки внутриклеточный домен интегрина связывается с цитоскелетом через адаптерные белки, такие как талин , α-актинин , филамин , винкулин и тензин . Многие др. Внутриклеточные сигнальные белки, такие как киназа фокальной адгезии , связываются и связываются с этим комплексом интегрин-адаптерный белок-цитоскелет, и это формирует основу фокальной адгезии.
Динамика адгезии с мигрирующими клетками [ править ]
Динамическая сборка и разборка фокальных адгезий играет центральную роль в миграции клеток . Во время миграции клеток изменяется как состав, так и морфология очаговой адгезии. Первоначально, малые (0.25μm²) фокальные спайки называемые координационные комплексы (FXS) образованы на переднем краю ячейки в ламеллиподий : они состоят из интегрина, и некоторые из адаптера белков, таких как Талин , паксиллина и тензина . Многие из этих фокальных комплексов не созревают и разбираются по мере удаления ламеллиподий. Однако некоторые фокальные комплексы созревают в более крупные и стабильные фокальные спайки и привлекают гораздо больше белков, таких как зиксин.. Привлечение компонентов к очаговой адгезии происходит упорядоченно, последовательно. [5] Оказавшись на месте, фокальная адгезия остается неподвижной по отношению к внеклеточному матриксу, и клетка использует его в качестве якоря, на котором она может подталкивать или тянуть себя через ECM. По мере того, как клетка продвигается по выбранному ею пути, данная фокальная адгезия перемещается все ближе и ближе к заднему краю клетки. На заднем крае клетки должна исчезнуть очаговая адгезия. Механизм этого плохо изучен и, вероятно, инициируется множеством различных методов в зависимости от обстоятельств клетки. Одна из возможностей состоит в том, что кальций-зависимая протеаза кальпаинучаствует: было показано, что ингибирование кальпаина приводит к ингибированию фокальной адгезии-ECM разделения. Компоненты фокальной адгезии относятся к числу известных субстратов кальпаина, и возможно, что кальпаин разрушает эти компоненты, чтобы помочь в разборке фокальной адгезии [6]
Ретроградный поток актина [ править ]
Сборка возникающих фокальных спаек сильно зависит от процесса ретроградного потока актина. Это явление в мигрирующей клетке, когда актиновые филаменты полимеризуются на переднем крае и текут обратно к телу клетки. Это источник тяги, необходимый для миграции; фокальная адгезия действует как молекулярная муфта, когда она привязывается к ЕСМ, и препятствует ретроградному движению актина, тем самым создавая тянущую (тяговую) силу в месте адгезии, которая необходима клетке для движения вперед. Это растяжение можно визуализировать с помощью микроскопии силы тяги.. Распространенная метафора для объяснения ретроградного потока актина - это большое количество людей, смываемых вниз по реке, и при этом некоторые из них цепляются за камни и ветви вдоль берега, чтобы остановить свое движение вниз по реке. Таким образом, на камень или ветку, за которые они держатся, создается тянущее усилие. Эти силы необходимы для успешной сборки, роста и созревания фокальных спаек. [7]
Природный биомеханический датчик [ править ]
Внеклеточные механические силы, которые проявляются через очаговые адгезии, могут активировать киназу Src и стимулировать рост спаек. Это указывает на то, что фокальные спайки могут функционировать как механические сенсоры, и предполагает, что сила, генерируемая миозиновыми волокнами, может способствовать созреванию фокальных комплексов. [8] Это получает дополнительную поддержку из того факта, что ингибирование сил, генерируемых миозином, ведет к медленной разборке фокальных адгезий за счет изменения кинетики оборота белков фокальной адгезии. [9]
Однако взаимосвязь между силами, воздействующими на фокальные спайки и их композиционное созревание, остается неясной. Например, предотвращение созревания фокальной адгезии путем ингибирования активности миозина или сборки стрессовых волокон не предотвращает сил, поддерживаемых фокальными адгезиями, и не предотвращает миграцию клеток. [10] [11] Таким образом, распространение силы через очаговые адгезии не может восприниматься клетками напрямую в любое время и в любых масштабах силы.
Их роль в механочувствительности важна для дуротаксиса .
См. Также [ править ]
- Актин
- TES (белок)
- Паксиллин
Ссылки [ править ]
- ^ Чен, CS; Алонсо, JL; Остуни, Э; Уайтсайдс, GM; Ingber, DE (2003). «Форма ячейки обеспечивает глобальный контроль сборки фокальной адгезии». Сообщения о биохимических и биофизических исследованиях . 307 (2): 355–61. DOI : 10.1016 / s0006-291x (03) 01165-3 . PMID 12859964 .
- ^ Зайдель-Бар, R; Коэн, М; Аддади, L; Гейгер, Б. (2004). «Иерархическая сборка адгезионных комплексов клеточного матрикса». Труды биохимического общества . 32 (3): 416–20. CiteSeerX 10.1.1.624.3354 . DOI : 10,1042 / bst0320416 . PMID 15157150 .
- ^ Замир, E; Гейгер, Б. (2001). «Молекулярная сложность и динамика клеточно-матричных адгезий». Журнал клеточной науки . 114 (20): 3583–90.
- ^ Ривелин, D; Замир, Э; Балабан, Северная Каролина; Шварц, США; Ишизаки, Т; Нарумия, S; Кам, З; Гейгер, Б; Бершадский, АД (2001). «Фокальные контакты как механодатчики: приложенная извне локальная механическая сила вызывает рост фокальных контактов с помощью mDia1-зависимого и ROCK-независимого механизма» . Журнал клеточной биологии . 153 (6): 1175–86. DOI : 10.1083 / jcb.153.6.1175 . PMC 2192034 . PMID 11402062 .
- ^ Зайдель-Бар, R; Коэн, М; Аддади, L; Гейгер, Б. (июнь 2004 г.). «Иерархическая сборка адгезионных комплексов клетка-матрица». Труды биохимического общества . 32 (Pt3): 416–20. CiteSeerX 10.1.1.624.3354 . DOI : 10,1042 / bst0320416 . PMID 15157150 .
- ^ Huttenlocher, A; Палецек, ИП; Лу, Q; Чжан, Вт; Меллгрен, Р.Л .; Lauffenburger, DA; Гинзберг, MH; Хорвиц, AF (1997). «Регулирование миграции клеток с помощью кальций-зависимой протеазы кальпаина» . Журнал биологической химии . 272 (52): 32719–22. DOI : 10.1074 / jbc.272.52.32719 . PMID 9407041 .
- ^ Гардель, ML; Sabass, B .; Ji, L .; Danuser, G .; Шварц, США; Уотерман, CM (8 декабря 2008 г.). «Растягивающее напряжение в очаговых спайках двухфазно коррелирует со скоростью ретроградного потока актина» . Журнал клеточной биологии . 183 (6): 999–1005. DOI : 10,1083 / jcb.200810060 . PMC 2600750 . PMID 19075110 .
- ^ Ван, Y; Botvinick, EL; Чжао, Y; Бернс, МВт; Усами, S; Tsien, RY; Чиен, С. (2005). «Визуализация механической активации Src» . Природа . 434 (7036): 1040–5. DOI : 10,1038 / природа03469 . PMID 15846350 .
- ^ Wolfenson, H .; Бершадский, А .; Хенис, Ю.И.; Гейгер, Б. (2011). «Напряжение, создаваемое актомиозином, контролирует молекулярную кинетику фокальных спаек» . J Cell Sci . 124 (9): 1425–32. DOI : 10,1242 / jcs.077388 . PMC 3078811 . PMID 21486952 .
- ^ Бенинго, Карен А .; Дембо, Мика; Каверина Ирина; Смолл, Дж. Виктор; Ван Юй-ли (2001-05-14). «Возникающие фокальные спайки несут ответственность за создание сильных движущих сил в мигрирующих фибробластах» . Журнал клеточной биологии . 153 (4): 881–888. DOI : 10.1083 / jcb.153.4.881 . ISSN 0021-9525 . PMC 2192381 . PMID 11352946 .
- ^ Стрикер, Джонатан; Аратин-Шаус, Ивонн; Оукс, Патрик В .; Гардель, Маргарет Л. (22.06.2011). «Пространственно-временные ограничения на силозависимый рост фокальных спаек» . Биофизический журнал . 100 (12): 2883–2893. DOI : 10.1016 / j.bpj.2011.05.023 . ISSN 1542-0086 . PMC 3123981 . PMID 21689521 .
Внешние ссылки [ править ]
- MBInfo - фокальная адгезия
- MBInfo - Узел фокальной адгезии
- MBInfo - Регулирование узла фокальной адгезии
- База данных AdhesomeFAnetwork со всеми известными белками фокальной адгезии и их биохимическими взаимодействиями
- Межклеточные связи
- Лаборатория клеточной адгезии Зайдель-Бар