Лисенин - это порообразующий токсин (PFT), присутствующий в целомической жидкости дождевого червя Eisenia fetida . Токсины, образующие поры, представляют собой группу белков, которые действуют как факторы вирулентности нескольких патогенных бактерий . Белки лисенина в основном участвуют в защите от эукариотических и прокариотических патогенов. [1] Следуя общему механизму действия PFT, лизенин выделяется как растворимый мономер, который специфически связывается с мембранным рецептором , сфингомиелином.в случае лизенина. После присоединения к мембране начинается олигомеризация, в результате чего поверх мембраны образуется нонамер, известный как препор. После конформационного изменения, которое может быть вызвано снижением pH , олигомер внедряется в мембрану в так называемом состоянии пор.
Мономер [ править ]
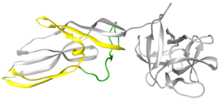
Лисенин - это белок, вырабатываемый целомоцитами - лейкоцитами дождевого червя Eisenia fetida . [2] Этот белок был впервые выделен из целомической жидкости в 1996 году и назван лизенином (от лизиса и Eisenia ). [3] Лисенин представляет собой относительно небольшую водорастворимую молекулу с молекулярной массой 33 кДа. С помощью рентгеновской кристаллографии лизенин был классифицирован как член семейства белков аэролизина по структуре и функциям. [4]Структурно каждый мономер лизенина состоит из рецепторного связывающего домена (серая глобулярная часть справа на Рисунке 1) и модуля порообразования (PFM); домены, общие для всего семейства аэролизинов. [4] Домен связывания рецептора лизенина имеет три мотива связывания сфингомиелина . Модуль формирования поры содержит области, которые претерпевают большие конформационные изменения, чтобы стать β-стволом в поре. [5]
Мембранные рецепторы [ править ]
Природной мембранной мишенью лизенина является липид плазматической мембраны животного, называемый сфингомиелином, расположенный в основном в его наружной створке, с участием по крайней мере трех из его фосфатидилхолиновых (PC) групп. [6] Сфингомиелин обычно связан с холестерином в липидных рафтах . [7] Холестерин, который усиливает олигомеризацию , обеспечивает стабильную платформу с высокой латеральной подвижностью, где встречи мономер-мономер более вероятны. [6] Было показано, что PFT способны реконструировать структуру мембраны, [8] иногда даже смешивая липидные фазы. [9]
Область β-цилиндра лисениновой поры, которая, как ожидается, будет погружена в гидрофобную область мембраны, является «детергентным поясом», областью высотой 3,2 нм, занятой детергентом в исследованиях поры с помощью криогенной электронной микроскопии (Cryo-EM). [10] С другой стороны, бислои сфингомиелина / холестерина имеют высоту около 4,5 нм. [11] Эта разница в высоте между лентой с моющим средством и бислоем сфингомиелина / холестерина подразумевает изгиб мембраны в области, окружающей пору, что называется отрицательным несоответствием. [12] Этот изгиб приводит к чистому притяжению между порами, которое вызывает их агрегацию.
Связывание, олигомеризация и вставка [ править ]
Связывание с мембраной является необходимым условием для инициации олигомеризации PFT. Мономеры лизенина специфически связываются со сфингомиелином через рецептор-связывающий домен. [13] Конечный олигомер лизенина состоит из девяти мономеров без количественных отклонений. [14] Когда мономеры лизенина связываются с областями мембраны, обогащенными сфингомиелином, они обеспечивают стабильную платформу с высокой латеральной подвижностью, что способствует олигомеризации. [15] Как и в случае с большинством PFT, олигомеризация лизенина происходит в двухступенчатом процессе, как было недавно показано.
Процесс начинается с того, что мономеры адсорбируются мембраной за счет определенных взаимодействий, что приводит к увеличению концентрации мономеров. Этому увеличению способствует небольшая площадь, где накапливается мембранный рецептор, в связи с тем, что большинство мембранных рецепторов PFT связаны с липидными рафтами. [16] Другим побочным эффектом, помимо увеличения концентрации мономера, является взаимодействие мономер-мономер. Это взаимодействие увеличивает олигомеризацию лизенина. После достижения критической пороговой концентрации одновременно образуются несколько олигомеров, хотя иногда они неполные. [17] В отличие от PFT из семейства холестерин-зависимых цитолизинов , [18] перехода от неполных олигомеров лизенина к полным олигомерам не наблюдалось.
Полная олигомеризация приводит к так называемому состоянию препоры - структуре на мембране. Определение структуры препора с помощью рентгеновских лучей или крио-ЭМ - сложный процесс, который до сих пор не дал никаких результатов. Единственная доступная информация о структуре препоры была предоставлена с помощью атомно-силовой микроскопии (АСМ). Измеренная высота препоры составляла 90 Å; и шириной 118 Å с внутренней порой 50 Å. [17] Модель препора была построена, выравнивая структуру мономера ( PDB : 3ZXD ) со структурой поры ( PDB : 5GAQ) Их рецептор-связывающими доменами (остатки от 160 до 297). Недавнее исследование аэролизина предполагает, что в соответствии с новыми доступными данными о введении аэролизина следует пересмотреть принятую в настоящее время модель для лисенинового препора. [19]
Конформационное изменение превращает PFM в трансмембранном & beta; баррель , что приводит к состоянию пор. [20] Триггерный механизм перехода из поры в поры в лизенине зависит от трех остатков глутаминовой кислоты (E92, E94 и E97) и активируется при снижении pH [21] от физиологических условий до достигаемых кислых условий. после эндоцитоза или увеличения внеклеточной концентрации кальция. [22] Эти три глутаминовые кислоты расположены в α-спирали, которая является частью PFM, а глутаминовые кислоты находятся в членах семейства аэролизинов в его PFM. Такое конформационное изменение приводит к уменьшению высоты олигомера на 2,5 нм согласно измерениям АСМ. [17]Основными размерами, используя рентгеновскую структуру пор лисенина, являются высота 97 Å, ширина 115 Å и внутренняя пора 30 Å. [20] Однако полная олигомеризация в нонамер не является обязательным условием для введения, поскольку могут быть обнаружены неполные олигомеры в состоянии пор. [17] Переход от препоры к поре может быть заблокирован в условиях тесноты, механизм, который может быть общим для всех β-PFT. Первый намек на эффект скопления при переходе от препоры к поре был дан в эффектах скопления в электрофизиологических экспериментах. [23] Высокоскоростные АСМ исследования инкубации лизенина на сфингомиелиновых / холестериновых мембранах показали, что в условиях тесноты переход от препоры к поре блокируется стерическими взаимодействиями. [24] [25] [26]
Последствия вставки [ править ]
Окончательные последствия образования пор под лизенином хорошо не задокументированы; однако считается, что он вызывает апоптоз с помощью трех возможных гипотез:
- Нарушение асимметрии сфингомиелина между двумя листочками липидного бислоя путем пробивания отверстий в мембране [27] и индукции липидного флип-флопа (переориентации липида с одного листочка мембранного бислоя на другой). [28]
- Повышение концентрации кальция в цитоплазме. [29]
- Снижение концентрации калия в цитоплазме. [30]
Биологическая роль [ править ]
Биологическая роль лизенина остается неизвестной. Было высказано предположение, что лизенин может играть роль защитного механизма против нападающих, таких как бактерии , грибы или мелкие беспозвоночные . [31] Однако активность лизенина зависит от связывания со сфингомиелином, которого нет в мембранах бактерий, грибов или большинства беспозвоночных. Скорее сфингомиелин в основном присутствует в плазматической мембране хордовых . [32] Другая гипотеза состоит в том, что дождевой червь, который способен выделять целомическую жидкость при стрессе, [33] [34] вызывает у своих позвоночных поведение избегания.хищники (например, птицы, ежи или кроты ). [35] Если это так, удаленный лизенин может быть более эффективным, если целомическая жидкость достигает глаза, где концентрация сфингомиелина в десять раз выше, чем в других органах тела. [36] Дополнительная гипотеза состоит в том, что резкий запах целомической жидкости, дающий дождевому червю его специфический эпитет foetida, является адаптацией против хищников . Однако остается неизвестным, способствует ли лизенин предотвращению хищников Eisenia . [37]
Приложения [ править ]
Электропроводящие свойства Лысенина изучаются годами. [38] Как и большинство порообразующих токсинов, лизенин образует неспецифический канал, проницаемый для ионов, небольших молекул и небольших пептидов. [39] Более трех десятилетий проводились исследования по поиску подходящих пор для преобразования в системы секвенирования нанопор , проводящие свойства которых можно регулировать посредством точечной мутации. [40] Из-за его аффинности связывания со сфингомиелином, лизенин (или просто рецептор-связывающий домен) использовался в качестве маркера флуоресценции для обнаружения сфингомиелинового домена в мембранах. [41]
Ссылки [ править ]
Эта статья была отправлена в WikiJournal of Science для внешнего научного рецензирования в 2019 году ( отчеты рецензентов ). Обновленный контент был повторно интегрирован на страницу Википедии по лицензии CC-BY-SA-3.0 ( 2019 ). Пересмотренная версия записи: Игнасио Л. Б. Мунгира; и другие. (17 августа 2019 г.). «Лысенин» (PDF) . WikiJournal of Science . 2 (1): 6. DOI : 10,15347 / WJS / 2019,006 . ISSN 2470-6345 . Викиданные Q76846397 .
- ^ Брун, Хайке; Винкельманн, Юлия; Андерсен, Кристиан; Андре, Йорг; Лейппе, Маттиас (2006). «Рассмотрение механизмов цитолитической и антибактериальной активности лизенина, защитного белка кольчатых червей Eisenia fetida» . Развитие и сравнительная иммунология . 30 (7): 597–606. DOI : 10.1016 / j.dci.2005.09.002 . PMID 16386304 .
- ^ Yilmaz, N .; Yamaji-Hasegawa, A .; Хуллин-Мацуда, Ф .; Кобаяши, Т. (2018). «Молекулярные механизмы действия сфингомиелин-специфического порообразующего токсина лизенина». Семинары по клеточной биологии и биологии развития . 73 : 188–198. DOI : 10.1016 / j.semcdb.2017.07.036 . PMID 28751253 .
- ^ Sekizawa, Y .; Hagiwara, K .; Накадзима, Т .; Кобаяши, Х. (1996). "Новый белок, лизенин, который вызывает сокращение изолированной аорты крысы: его очистка от целомической жидкости дождевого червя, Eisenia foetida " . Биомедицинские исследования . 17 (3): 197–203. DOI : 10.2220 / biomedres.17.197 .
- ^ a b De Colibus, L .; Sonnen, AF-P .; Моррис, KJ; Зиберт, Калифорния; Abrusci, P .; Plitzko, J .; Ходник, В .; Leippe, M .; Volpi, E .; Anderluh, G .; Гилберт, RJC (2012). «Структуры Лисенина обнаруживают общее эволюционное происхождение порообразующих белков и его способ распознавания сфингомиелина» . Структура . 20 (9): 1498–1507. DOI : 10.1016 / j.str.2012.06.011 . PMC 3526787 . PMID 22819216 .
- ^ Бокори-Браун, М .; Мартин, Т.Г.; Нейлор, CE; Басак, АК; Титболл, RW; Савва, CG (2016). «Крио-ЭМ структура поры лизенина объясняет встраивание в мембрану белков семейства аэролизинов» . Nature Communications . 7 (1): 11293. Bibcode : 2016NatCo ... 711293B . DOI : 10.1038 / ncomms11293 . PMC 4823867 . PMID 27048994 .
- ^ a b Ishitsuka, R .; Кобаяши, Т. (2007). «Холестерин и соотношение липид / белок контролируют олигомеризацию сфингомиелин-специфического токсина, лисенина» . Биохимия . 46 (6): 1495–1502. DOI : 10.1021 / bi061290k . PMID 17243772 . S2CID 22016219 .
- ^ Саймонс, К .; Герл, MJ (2010). «Оживление мембранных плотов: новые инструменты и идеи». Обзоры природы Молекулярная клеточная биология . 11 (10): 688–699. DOI : 10.1038 / nrm2977 . PMID 20861879 . S2CID 1866391 .
- ^ Ros, U .; Гарсиа-Саес, AJ (2015). «Больше, чем поры: взаимодействие порообразующих белков и липидных мембран». Журнал мембранной биологии . 248 (3): 545–561. DOI : 10.1007 / s00232-015-9820-у . PMID 26087906 . S2CID 16305100 .
- ^ Yilmaz, N .; Кобаяши, Т. (2015). «Визуализация реорганизации липидной мембраны, вызванной порообразующим токсином с использованием высокоскоростной атомно-силовой микроскопии». САУ Нано . 9 (8): 7960–7967. DOI : 10.1021 / acsnano.5b01041 . PMID 26222645 .
- ^ Бокори-Браун, М .; Мартин, Т.Г.; Нейлор, CE; Басак, АК; Титболл, RW; Савва, CG (2016). «Крио-ЭМ структура поры лизенина объясняет встраивание в мембрану белков семейства аэролизинов» . Nature Communications . 7 (1): 11293. Bibcode : 2016NatCo ... 711293B . DOI : 10.1038 / ncomms11293 . PMC 4823867 . PMID 27048994 .
- Перейти ↑ Quinn, PJ (2013). «Структура бислоев сфингомиелина и комплексов с холестерин образующими мембранными рафтами». Ленгмюра . 29 (30): 9447–9456. DOI : 10.1021 / la4018129 . PMID 23863113 .
- ^ Guigas, G .; Вайс, М. (2016). «Влияние белкового краудинга на мембранные системы» . Biochimica et Biophysica Acta (BBA) - Биомембраны . 1858 (10): 2441–2450. DOI : 10.1016 / j.bbamem.2015.12.021 . PMID 26724385 .
- ^ De Colibus, L .; Sonnen, AF-P .; Моррис, KJ; Зиберт, Калифорния; Abrusci, P .; Plitzko, J .; Ходник, В .; Leippe, M .; Volpi, E .; Anderluh, G .; Гилберт, RJC (2012). «Структуры Лисенина обнаруживают общее эволюционное происхождение порообразующих белков и его способ распознавания сфингомиелина» . Структура . 20 (9): 1498–1507. DOI : 10.1016 / j.str.2012.06.011 . PMC 3526787 . PMID 22819216 .
- ^ Munguira, I .; Casuso, I .; Takahashi, H .; Rico, F .; Мияги, А .; Chami, M .; Шейринг, С. (2016). «Стекловидная мембрана диффузии белков в переполненной мембране» (PDF) . САУ Нано . 10 (2): 2584–2590. DOI : 10.1021 / acsnano.5b07595 . PMID 26859708 .
- ^ Ishitsuka, R .; Кобаяши, Т. (2007). «Холестерин и соотношение липид / белок контролируют олигомеризацию сфингомиелин-специфического токсина, лизенина» . Биохимия . 46 (6): 1495–1502. DOI : 10.1021 / bi061290k . PMID 17243772 . S2CID 22016219 .
- ^ Lafont, F .; Ван Дер Гут, ФГ (2005). «Бактериальная инвазия через липидные рафты» . Клеточная микробиология . 7 (5): 613–620. DOI : 10.1111 / j.1462-5822.2005.00515.x . PMID 15839890 . S2CID 26547616 .
- ^ a b c d Yilmaz, N .; Yamada, T .; Greimel, P .; Uchihashi, T .; Андо, Т .; Кобаяши, Т. (2013). «Визуализация в реальном времени сборки специфического для сфингомиелина токсина на плоских липидных мембранах» . Биофизический журнал . 105 (6): 1397–1405. Bibcode : 2013BpJ ... 105.1397Y . DOI : 10.1016 / j.bpj.2013.07.052 . PMC 3785888 . PMID 24047991 .
- ^ Mulvihill, E .; van Pee, K .; Mari, SA; Мюллер, диджей; Йылдыз, Ö. (2015). «Непосредственное наблюдение липид-зависимого механизма самосборки и порообразования цитолитического токсина Listeriolysin O». Нано-буквы . 15 (10): 6965–6973. Bibcode : 2015NanoL..15.6965M . DOI : 10.1021 / acs.nanolett.5b02963 . PMID 26302195 .
- ^ Iacovache, Иоан; Де Карло, Саша; Чирауки, Нурия; Даль Пераро, Маттео; ван дер Гут, Ф. Гису; Зубер, Бенуа (2016). «Крио-ЭМ структура вариантов аэролизина выявляет новую белковую складку и процесс порообразования» . Nature Communications . 7 : 12062. Bibcode : 2016NatCo ... 712062I . DOI : 10.1038 / ncomms12062 . PMC 4947156 . PMID 27405240 .
- ^ a b Бокори-Браун, М .; Мартин, Т.Г.; Нейлор, CE; Басак, АК; Титболл, RW; Савва, CG (2016). «Крио-ЭМ структура поры лизенина объясняет встраивание в мембрану белков семейства аэролизинов» . Nature Communications . 7 (1): 11293. Bibcode : 2016NatCo ... 711293B . DOI : 10.1038 / ncomms11293 . PMC 4823867 . PMID 27048994 .
- ^ Munguira, ILB; Takahashi, H .; Casuso, I .; Шойринг, С. (2017). «Введение в мембрану лизенинового токсина зависит от pH, но не зависит от соседних лизенинов» . Биофизический журнал . 113 (9): 2029–2036. Bibcode : 2017BpJ ... 113.2029M . DOI : 10.1016 / j.bpj.2017.08.056 . PMC 5685674 . PMID 29117526 .
- ^ Munguira, ILB (2019). «Механизм введения токсина лизенина зависит от кальция» . bioRxiv . DOI : 10.1101 / 771725 .
- ^ Krueger, E .; Bryant, S .; Shrestha, N .; Clark, T .; Hanna, C .; Pink, D .; Фологея, Д. (2015). «Эффекты внутримембранного затора на индуцированном напряжением стробировании канала лизенина» . Европейский биофизический журнал . 45 (2): 187–194. DOI : 10.1007 / s00249-015-1104-Z . PMC 4803513 . PMID 26695013 .
- ^ Munguira, ILB (2017). Эффект скученности в жизненном цикле Лисенина, изученный с помощью высокоскоростной атомно-силовой микроскопии (PhD). Университет Экс-Марсель.
- ^ Munguira, NL (2020). «Активность порообразующего токсина лизенина регулируется скученностью». Нанотехнологии . DOI : 10.1088 / 1361-6528 .
- ^ Munguira, ILB (2020). «Стерическая блокада лизенинового токсина скученностью». bioRxiv . DOI : 10.1101 / 2020.05.02.073940 .
- ^ Грин, DR (2000). «Апоптоз и гидролиз сфингомиелина» . Журнал клеточной биологии . 150 (1): F5 – F8. DOI : 10.1083 / jcb.150.1.F5 . PMC 2185551 . PMID 10893276 .
- ^ Ros, U .; Гарсиа-Саес, AJ (2015). «Больше, чем поры: взаимодействие порообразующих белков и липидных мембран». Журнал мембранной биологии . 248 (3): 545–561. DOI : 10.1007 / s00232-015-9820-у . PMID 26087906 . S2CID 16305100 .
- ^ Оррениус, S .; Животовский, Б .; Никотера, П. (2003). «Регулирование гибели клеток: связь кальций – апоптоз». Обзоры природы Молекулярная клеточная биология . 4 (7): 552–565. DOI : 10.1038 / nrm1150 . PMID 12838338 . S2CID 19079491 .
- ↑ Yu, SP (2003). «Регуляция и критическая роль гомеостаза калия в апоптозе». Прогресс нейробиологии . 70 (4): 363–386. DOI : 10.1016 / s0301-0082 (03) 00090-х . PMID 12963093 . S2CID 13893235 .
- ^ Ballarin, L .; Каммарата, М. (2016). Уроки иммунитета: от одноклеточных организмов до млекопитающих . Академическая пресса. ISBN 9780128032527.
- ^ Кобаяши, H .; Sekizawa, Y .; Aizu, M .; Умеда, М. (2000). «Летальные и нелетальные реакции сперматозоидов различных позвоночных и беспозвоночных на лизенин, белок из целомической жидкости дождевого червя Eisenia foetida ». Журнал экспериментальной зоологии . 286 (5): 538–549. doi : 10.1002 / (sici) 1097-010x (20000401) 286: 5 <538 :: aid-jez12> 3.0.co; 2-w . PMID 10684578 .
- ^ Sukumwang, N .; Умедзава, К. (2013). "Порообразующий токсин Лисенин, полученный из дождевых червей, и скрининг его ингибиторов" . Токсины . 5 (8): 1392–1401. DOI : 10,3390 / toxins5081392 . PMC 3760042 . PMID 23965430 .
- ^ Кобаяши, H .; Ohta, N .; Умеда, М. (2004). «Биология лизенина, белка целомической жидкости дождевого червя Eisenia foetida ». Международный обзор цитологии . 236 : 45–99. DOI : 10.1016 / S0074-7696 (04) 36002-X . ISBN 9780123646408. PMID 15261736 .
- ^ Swiderska, B .; Кедрацка-Крок, С .; Panz, T .; Морган, AJ; Фалниовский, А .; Grzmil, P .; Плытыч, Б. (2017). «Белки семейства Лысенина в целомоцитах дождевых червей - Сравнительный подход». Развитие и сравнительная иммунология . 67 : 404–412. DOI : 10.1016 / j.dci.2016.08.011 . PMID 27567602 . S2CID 19895826 .
- Перейти ↑ Berman, ER (1991). Биохимия глаза . Springer. DOI : 10.1007 / 978-1-4757-9441-0 . ISBN 978-1-4757-9441-0. S2CID 41192657 .
- ^ Эдвардс, Калифорния; Болен, П.Дж. (1996). Биология и экология дождевых червей . Springer Science & Business Media. ISBN 978-0-412-56160-3.
- ^ Брайант, S .; Clark, T .; Thomas, C .; Посуда, К .; Bogard, A .; Calzacorta, C .; Prather, D .; Фологея, Д. (2018). «Понимание механизма регулирования напряжения порообразующего токсина Лисенина» . Токсины . 10 (8): 334. DOI : 10,3390 / toxins10080334 . PMC 6115918 . PMID 30126104 .
- ^ Shrestha, N .; Брайант, SL; Thomas, C .; Richtsmeier, D .; Pu, X .; Tinker, J .; Фологея, Д. (2017). «Стохастическое зондирование ангиотензина II с лизениновыми каналами» . Научные отчеты . 7 (1): 2448. Bibcode : 2017NatSR ... 7.2448S . DOI : 10.1038 / s41598-017-02438-0 . PMC 5446423 . PMID 28550293 .
- ^ Deamer, D .; Akeson, M .; Брантон, Д. (2016). «Три десятилетия секвенирования нанопор» . Природа Биотехнологии . 34 (5): 518–524. DOI : 10.1038 / nbt.3423 . PMC 6733523 . PMID 27153285 .
- ^ Ishitsuka, R .; Кобаяши, Т. (2004). «Лысенин: новый инструмент для исследования липидной организации мембран». Anatomical Science International . 79 (4): 184–190. DOI : 10.1111 / j.1447-073x.2004.00086.x . PMID 15633456 . S2CID 1558393 .
Внешние ссылки [ править ]
- https://www.theses.fr/2017AIXM0124