| Немертя | |
|---|---|
 | |
| Научная классификация | |
| Королевство: | Animalia |
| Подцарство: | Eumetazoa |
| Клэйд : | ParaHoxozoa |
| Клэйд : | Bilateria |
| Клэйд : | Нефрозоа |
| (без рейтинга): | Протостомия |
| (без рейтинга): | Спиралия |
| Суперфилум: | Лофотрохозоа |
| Тип: | Немертя Шульце , 1851 г. |
| Классы | |
| Синонимы [1] | |
Nemertini | |
{kind=link}
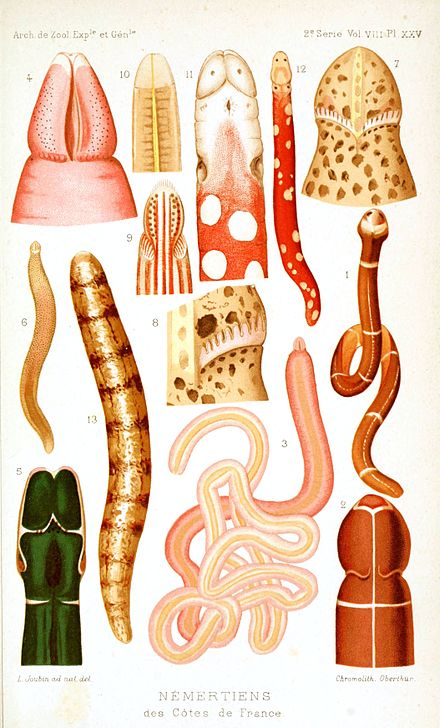
Немертин являются филюмом из беспозвоночных животных , также известные как ленточных черви или Хоботковых червь . [2] Альтернативные названия филума включали Nemertini , Nemertinea и Rhynchocoela . [1] Большинство из них очень тонкие, обычно всего несколько миллиметров в ширину, хотя некоторые имеют относительно короткие, но широкие тела. Многие имеют узоры желтого, оранжевого, красного и зеленого цветов. Передняя кишка, желудок и кишечник проходят немного ниже средней линии тела, анус находится на кончике хвоста, а рот - под передней частью. Чуть выше кишечника находитсяринхоцель - полость, которая в основном проходит над средней линией и заканчивается немного ниже задней части тела. У всех видов есть хоботок, который находится в ринхоцеле в неактивном состоянии, но выворачивается (выворачивается наизнанку), выходит прямо над ртом и захватывает добычу с ядом. Сильно растяжимая мышца в задней части ринхоцеля втягивает хоботок, когда приступ заканчивается. Некоторые виды с короткими телами фильтруют корм и имеют присоски на переднем и заднем концах, с помощью которых они прикрепляются к хозяину .
Мозг представляет собой кольцо из четырех ганглиев , расположенных вокруг ринхоцеля возле его переднего конца. По крайней мере, пара брюшных нервных тяжей соединяется с мозгом и проходит по длине тела. У большинства немертин есть различные хеморецепторы , а на головах некоторых видов есть несколько глазков пигментных чашечек . Эти глазки могут обнаруживать свет, но не могут формировать изображение. Немертин дышит через кожу. У них есть по крайней мере два боковых сосуда, которые соединяются на концах в петлю, и они вместе с ринхоцелем заполнены жидкостью. Сердца нет, а поток жидкости зависит от сокращения мышц сосудов и стенок тела. Чтобы отфильтровать растворимые отходы,Ячейки пламени встроены в переднюю часть двух боковых резервуаров для жидкости и удаляют отходы через сеть труб наружу.
Все немертины передвигаются медленно, используя свои внешние реснички, чтобы скользить по поверхности по следу слизи , в то время как более крупные виды используют мышечные волны, чтобы ползать, а некоторые плавают по дорсо-вентральным волнам . Некоторые живут в открытом океане, а остальные находят или прячутся на дне. Около десятка видов обитают в пресной воде, в основном в тропиках и субтропиках, а еще десяток видов обитает на суше в прохладных влажных местах. Большинство немертин - плотоядные животные , питающиеся кольчатыми червями , моллюсками и ракообразными . Несколько видов падальщики , и несколько видов живут commensally внутри мантийной полостимоллюски . Некоторые виды опустошили коммерческий лов моллюсков и крабов. У немертинов мало хищников. Два вида продаются в качестве приманки для рыбы.
У большинства видов полы раздельны, но все пресноводные виды гермафродиты . Немертины часто имеют многочисленные временные гонады ( яичники или семенники ) и строят временные гоноды (протоки, из которых выходят яйцеклетки или сперматозоиды), открывающиеся к гонопорам , по одному на гонаду, когда яйцеклетки и сперматозоиды готовы. Яйца обычно оплодотворяются внешне. Некоторые виды сбрасывают их в воду, а другие различными способами защищают свои яйца. Оплодотворенная яйцеклетка делится путем спирального дробления и растет путем детерминированного развития , при котором судьбу клетки обычно можно предсказать по ее предшественникам в процессе деления. Эмбрионы большинстваТаксоны развиваются либо непосредственно с образованием молоди (как и взрослые особи, но меньшего размера), либо с образованием плоских личинок, у которых длинная ось личинки совпадает с осью молоди. Однако некоторые из них образуют личинку пилидиума , в которой у развивающейся молоди есть кишка, расположенная поперек тела личинки, и обычно она поедает остатки личинки, когда она появляется. Тела некоторых видов легко фрагментируются, и даже части около хвоста могут вырасти полностью.
Было высказано предположение, что три ископаемых вида могут быть немертами, но ни один из них не подтвержден. Традиционная таксономия делит тип на два класса : Anopla («невооруженные» - их хоботки не имеют маленького кинжала) с двумя отрядами и Enopla («вооруженные» кинжалом) с двумя отрядами. Однако сейчас принято считать, что Anopla являются парафилетическими (дали начало другой группе), поскольку один отряд более тесно связан с Enopla, чем с другим отрядом Anopla. Тип Nemertea монофилетический . Его синапоморфиивключают ринхоцель и разворотный хоботок. Традиционная систематика гласит, что немертины тесно связаны с плоскими червями . Оба типа считаются членами Lophotrochozoa , очень большого «супертипа», который также включает моллюсков , кольчатых червей , брахиопод , мшанок и многих других протостомов .
История [ править ]
В 1555 году Олаус Магнус написал о морском червяке, который, по-видимому, был 17,76 метра (58,3 фута) в длину («40 локтей»), примерно ширины руки ребенка, и от прикосновения которого рука опухла. Уильям Борлас написал в 1758 году «морского длинного червя», а в 1770 году Гуннерус написал формальное описание этого животного, которое он назвал Ascaris longissima . Его нынешнее название Lineus longissimus впервые было использовано в 1806 году Сауерби. [3] В 1995 г. было описано 1149 видов, сгруппированных в 250 родов. [4]
Немертея названа в честь греческой морской нимфы Немертес, одной из дочерей Нерея и Дориды . [5] Альтернативные названия филума включали Nemertini , Nemertinea и Rhynchocoela . [1] Nemertodermatida это отдельная фила, чьи близкие родственники по всей видимости, является бескишечные турбеллярии . [6] [7]
Описание [ править ]
Строение тела и основные полости [ править ]
Немертины - очень необычные животные. [8]
Типичное тело немертины очень стройное по сравнению с длиной. [9] Самые маленькие - несколько миллиметров в длину, [10] большинство - менее 20 сантиметров (7,9 дюйма), а некоторые превышают 1 метр (3,3 фута). Самый длинный животное когда - либо найденных, на 54 метров (177 футов) в длину, может представлять собой образец Lineus длиннейшей , [9] [Примечание 1] , хотя L. длиннейшей обычно составляет всего несколько миллиметров в ширину. [8] Тела большинства немертинов могут сильно растягиваться, до 10 раз превышающей длину покоя у некоторых видов, [8] [9], но при потревожении сокращаются до 50% и увеличиваются в ширину до 300%. [12]Некоторые из них имеют относительно короткие, но широкие тела, например, Malacobdella Grossa имеет длину до 3,5 см (1,4 дюйма) и ширину 1 см (0,39 дюйма) [9] [17], а некоторые из них гораздо менее эластичны. [8] Более мелкие немертины имеют приблизительно цилиндрическую форму, но более крупные виды уплощены дорсо-вентрально . Многие из них имеют видимые узоры в различных сочетаниях желтого, оранжевого, красного и зеленого цветов. [9]
Наружный слой тела не имеет кутикулу , но состоит из ресничного и железистого эпителия , содержащего rhabdites , [10] , которые образуют слизь , в которой реснички глиссады. [18] Каждая ресничная клетка имеет множество ресничек и микроворсинок . [9] Самый внешний слой лежит на утолщенной базальной мембране , дерме . [10] Рядом с дермой есть по крайней мере три слоя мышц, некоторые из которых являются круглыми, а некоторые - продольными. [9] Комбинации типов мышц различаются в зависимости от класса., но они не связаны с различиями в движении. [10] Немертины также имеют дорсо-вентральные мышцы, которые сглаживают животных, особенно у более крупных видов. [9] Внутри концентрических трубок этих слоев находится мезенхима , своего рода соединительная ткань . [10] У пелагических видов эта ткань студенистая и плавучая. [9]
Рот вентральный и немного позади передней части тела. Передняя кишка, желудок и кишечник проходят немного ниже средней линии тела, а задний проход находится на кончике хвоста. [19] Над кишечником и отделен от кишечника мезенхимой, находится ринхоцель , полость, которая в основном проходит выше средней линии и заканчивается немного ниже задней части тела. Ринхоцель класса Anopla имеет отверстие немного ближе к передней части рта, но все же находится под передней частью тела. В другом классе, Enopla , рот и передняя часть ринхоцеля имеют общее отверстие. [9] rhynchocoel является целомом , так как она выстлана эпителием .[10]
Хоботок и кормление [ править ]
Хобот является клубящийся стенки тела, и сидит в rhynchocoel , когда неактивен. [10] Когда мышцы в стенке ринхоцеля сжимают жидкость в ринхоцеле, давление заставляет хоботок прыгать наизнанку, чтобы атаковать добычу животного по каналу, называемому ринходом, и через отверстие - поре хоботка. Хоботок имеет мышцу, которая прикрепляется к задней части ринхоцеля и может растягиваться до 30 раз своей неактивной длины, а затем втягивать хоботок. [9]
Хоботок класса Anopla («невооруженный» [20] ) выходит из отверстия, которое отделено от рта [9], обвивается вокруг добычи и обездвиживает ее липкими ядовитыми выделениями. [19] Anopla может атаковать, как только они попадают в диапазон хоботка. [21] У некоторых Anopla есть разветвленные хоботки, которые можно описать как «массу липких спагетти». [9] Затем животное затягивает добычу в пасть. [10]
У большинства представителей класса Enopla («вооруженный» [20] ) хоботок выходит из общего отверстия ринхоцеля и рта. Типичный представитель этого класса имеет стилет , известковый заусенец, [9] которым животное много раз наносит удар жертве, чтобы ввести токсины и пищеварительные выделения. Затем добычу проглатывают целиком или, после частичного переваривания, ее ткани засасывают в рот. [19] Стилет прикреплен примерно на одной трети расстояния от конца вывернутого хоботка, который выдается только на то, чтобы обнажить стилет. По обе стороны от активного стилета находятся мешочки, содержащие запасные стилеты, заменяющие активный стилет по мере роста животного или потери активного.[9] Вместо одного стилета у Polystilifera есть подушечка с множеством крошечных стилетов, и у этих животных есть отдельные отверстия для хоботка и рта, в отличие от других Enopla. [20] [22] Энопла может атаковать только после контакта с добычей. [21]
Некоторые немертинцы, такие как L. longissimus , поглощают органическую пищу в растворе через кожу, что может сделать их длинные и стройные тела преимуществом. [8] Подвеска кормления встречается только среди специализированных симбиотических bdellonemerteans , [21] , которые имеют хобот , но не стилет, а также использовать присоски для крепления себя двустворчатых . [23]
Система дыхания и кровообращения [ править ]
У немертинов нет специализированных жабр , и дыхание происходит по поверхности тела, которая длинная, а иногда и приплюснутая. Как и другие животные с толстыми стенками тела, они используют циркуляцию жидкости, а не диффузию, для перемещения веществ через свое тело. Система кровообращения состоит из ринхоцеля и периферических сосудов [24], в то время как их кровь находится в основной полости тела. [25] Жидкость в ринхоцеле перемещает вещества в хоботок и из него и действует как жидкий скелет, вывертывая хоботок и роя норы. Сосуды обеспечивают циркуляцию жидкости по всему телу, а ринхоцель обеспечивает собственное местное кровообращение.[24] Кровеносные сосуды представляют собой систему целомов. [26]
В простейшем типе кровеносной системы два боковых сосуда соединяются на концах, образуя петлю. Однако у многих видов есть дополнительные продольные и поперечные сосуды. Нет ни сердца, ни перекачивающих сосудов [27], а поток жидкости зависит от сокращения сосудов и мышц стенок тела. У некоторых видов кровообращение прерывистое, и жидкость приливает и течет по длинным сосудам. [24] Жидкость в сосудах обычно бесцветная, но у некоторых видов она содержит клетки желтого, оранжевого, зеленого или красного цвета. Красный тип содержит гемоглобин и переносит кислород, но функция других пигментов неизвестна. [24]
Экскреция [ править ]
Nemertea использует органы, называемые протонефридиями [24], для вывода растворимых отходов жизнедеятельности, особенно азотистых побочных продуктов клеточного метаболизма . [28] У немертиновых протонефридий клетки пламени, которые отфильтровывают отходы, встроены в переднюю часть двух боковых сосудов с жидкостью. Ячейки пламени удаляют отходы в два сборных канала, по одному с каждой стороны, и каждый канал имеет одну или несколько нефридиопор, через которые отходы выходят. Полукоземные и пресноводные немертины имеют гораздо больше ячеек пламени, чем морские пехотинцы, иногда тысячи. Причина может заключаться в том, что осморегуляция затруднена в неморской среде. [24]
Нервная система и чувства [ править ]
Центральная нервная система состоит из головного мозга и парных вентральных нервных шнуров , которые подключаются к мозгу и бегам по длине тела. Мозг представляет собой кольцо из четырех ганглиев , масс нервных клеток, расположенных вокруг rhynchocoel вблизи ее переднего конца [29] - в то время как мозг большинство первичноротых беспозвоночных облегать кишки. [30] Большинство видов немертин имеют только одну пару нервных связок, многие виды имеют дополнительные парные связки, а некоторые виды также имеют спинной канатик. [29] У некоторых видов связки лежат внутри кожи, но у большинства они более глубокие, внутри мышечных слоев.[31] Центральная нервная система часто бывает красной или розовой, потому что она содержит гемоглобин . Это сохраняет кислород для пиковой активности или когда животное испытывает кислородное голодание , например, зарываясь в бескислородные отложения . [29]
У некоторых видов есть парные церебральные органы - мешочки, единственные отверстия которых выходят наружу. У других видов на передней части головы есть непарные вывернутые органы. У некоторых есть прорези по бокам головы или бороздки наискосок через голову, и они могут быть связаны с парными церебральными органами. Считается, что все они являются хеморецепторами , а органы головного мозга также могут способствовать осморегуляции . Маленькие ямки в эпидермисе кажутся сенсорами. [29] На голове некоторых видов есть несколько глазков пигментных чашечек , [29] которые могут обнаруживать свет, но не формировать изображение. [32] У большинства немертид от двух до шести глазков, хотя у некоторых их сотни. [31]Несколько крошечных видов , которые живут между зернами песка имеют Статоцисты , [29] , смысл баланса. [33]
Paranemertes peregrina , который питается полихетами, может следовать по слизистым следам жертвы и находить ее нору, возвращаясь назад по своему собственному следу слизи. [19]
Движение [ править ]
Немертины обычно передвигаются медленно [10], хотя иногда задокументировано, что они успешно охотятся на пауков или насекомых. [34] Большинство немертинов используют свои внешние реснички, чтобы скользить по поверхностям по следу слизи , часть которой производится железами в голове. Более крупные виды используют мышечные волны для ползания, а некоторые водные виды плавают дорсо-вентральными волнами. Некоторые виды роют норы посредством мышечной перистальтики и обладают мощными мускулами. [9] Некоторые виды подотряда Monostilifera , хоботок которых имеет один активный стилет, передвигаются, вытягивая хоботок, прикрепляя его к объекту и притягивая животное к объекту. [20]
Размножение и жизненный цикл [ править ]
Более крупные виды часто распадаются при стимуляции, а фрагменты часто превращаются в полные особи. Некоторые виды регулярно фрагментируются, и даже части около хвоста могут вырасти полностью. [35] Все размножаются половым путем , и большинство видов гонохорические (полы раздельные), [10] [35], но все пресноводные формы являются гермафродитами . [25]
Немертины часто имеют многочисленные временные гонады ( яичники или семенники ), образующие ряды вниз по каждой стороне тела в мезенхиме . [25] [35] Временные гоноды (каналы, из которых выходят яйцеклетки или сперматозоиды [36] ), по одному на гонаду, строятся, когда яйцеклетка и сперма готовы. [35] Яйца, как правило, оплодотворяются извне. Некоторые виды сбрасывают их в воду, некоторые кладут в нору или трубку, а некоторые защищают их коконами или студенистыми нитями. [35] Некоторые батипелагические(глубоководные) виды имеют внутреннее оплодотворение , и некоторые из них живородящие , выращивая свои эмбрионы в теле самки. [25] [35]
В зиготы (оплодотворенной яйцеклетки) делит по спирали расщепления и растет детерминированным развитием , [35] , в котором судьба клетки обычно может быть предсказана от своих предшественников в процессе разделения. [8] Эмбрионы большинства таксонов развиваются либо непосредственно с образованием подростков (как взрослые , но меньше) или с образованием planuliform личинок , [35] , в котором длинная оси личинки является таким же , как несовершеннолетний. [30] Стадия плоской личинки может быть недолгой и лецитотрофной («желточный») до того, как стать молодым, [35] или может бытьпланктотрофный , некоторое время плавает и поедает добычу крупнее микроскопических частиц. [30] Тем не менее, многие члены ордена Heteronemertea и palaeonemertean семейства Hubrechtidae образует pilidium личинку, которая может захватить одноклеточные водоросли и которые Маслаков описывает как как войлочный колпачок с ухом закрылки снесены. У него есть кишка, проходящая поперек тела, рот между «ушными раковинами», но нет заднего прохода. Образуется небольшое количество имагинальных дисков , окружающих архентерон.(развитие кишечника) и сливаются, образуя молодь. Когда он полностью сформирован, детеныш вырывается из тела личинки и обычно съедает его во время этого катастрофического метаморфоза . [30] Эта личиночная стадия уникальна тем, что в процессе развития не участвуют Hox-гены , которые обнаруживаются только у молоди, развивающейся внутри личинок. [37]
Сообщается, что продолжительность жизни вида Paranemertes peregrina составляет около 18 месяцев. [31]
Экологическое значение [ править ]
Большинство немертины морские животные , которые Нора в отложениях, скрываются в щелях между оболочками, камнями или holdfasts из водорослей или сидячих животных. Некоторые живут глубоко в открытом океане и имеют студенистые тела. Другие строят полупостоянные норы, выстланные слизью, или производят целлофановые трубки. В основном в тропиках и субтропиках около 12 видов встречаются в пресной воде [9], и около десятка видов обитают на суше в прохладных и влажных местах, например, под гниющими бревнами. [8]
Наземный Argonemertes dendyi является уроженцем Австралии, но был обнаружен на Британских островах , в Сан-Мигеле на Азорских островах , на Гран-Канарии и в лавовой трубе в Каумане на острове Гавайи . Он может строить кокон, что позволяет ему избежать высыхания во время транспортировки, и он может быстро создавать популяции в новых районах, поскольку это протандрозный гермафродит . [38] Другой наземный род , Geonemertes , в основном встречается в Австралазии.но есть виды на Сейшельских островах , широко в Индо-Тихоокеанском регионе , на Тристан-да-Кунья в Южной Атлантике, во Франкфурте , на Канарских островах , на Мадейре и на Азорских островах. [4] Geonemertes pelaensis был причастен к сокращению численности местных видов членистоногих на островах Огасавара , где он был завезен в 1980-е годы. [39]
Большинство из них являются плотоядные , питающиеся аннелид , моллюсками и ракообразными , [19] и может убить кольчатых червей из об их собственного размера. Иногда ловят рыбу, как живую, так и мертвую. Насекомые и многоножки - единственная известная жертва двух наземных видов Argonemertes . [21] Некоторые немертины - падальщики , [19] и они, как правило, обладают хорошей хеморецепцией на расстоянии («запахом») и не избирательны в отношении своей добычи. [21] Некоторые виды живут комменсально внутри мантии.полости моллюсков и питаются микроорганизмами, отфильтрованными хозяином. [40]
Около Сан-Франциско немертин Carcinonemertes errans съел около 55% от общего производства яиц своего хозяина, краба Dungeness Metacarcinus magister . C. errans считается значительным фактором в упадке промысла крабов-навозников. [21] Другие прибрежные немертины разрушили русла моллюсков . [9]
Немногочисленные хищники немертин включают донных рыб, некоторых морских птиц, некоторых беспозвоночных, включая подковообразных крабов , и других немертин. [9] Кожа немертин выделяет токсины, отпугивающие многих хищников, но некоторые крабы могут очистить немертинов одним когтем, прежде чем съесть их. [25] Американский Cerebratulus lacteus и южноафриканский Polybrachiorhynchus dayi , оба называемые «ленточными червями» в соответствующих местах обитания , продаются в качестве приманки для рыбы. [9]
Таксономия [ править ]
- Класс Anopla («безоружный»). Включает животных с хоботком без стилета и ртом под и позади мозга. [20]
- Отряд Palaeonemertea . Насчитывает 100 морских видов. Стенка их тела имеет наружные круговые и внутренние продольные мышцы. Кроме того, Carinoma tremaphoros имеет круговые и продольные внутренние мышцы эпидермиса ; дополнительные мышечные слои, по-видимому, необходимы для прохождения через перистальтику . [20]
- Заказать Heteronemertea . Насчитывает около 400 видов. Большинство из них морские, но три - пресноводные. Их мышцы стенки тела расположены в четыре слоя, поочередно круговые и продольные, начиная с самого внешнего слоя. В отряд входят сильнейшие пловцы. У двух родов хоботки разветвленные. [20]
- Класс Enopla («вооруженный»). У всех есть стилеты, кроме отряда Bdellonemertea . Их рот расположен под мозгом и впереди него. Их основные нервные связки проходят внутри мышц стенок тела. [20]
- Заказать Бделлонемертя. Включает семь видов, шесть из которых живут как комменсалы в мантии крупных моллюсков и один - в мантии пресноводной улитки. Хозяева фильтруют корм, а все хозяева крадут у них еду. Эти немертины имеют короткие и широкие тела и не имеют стилетов, но имеют сосательный зев и задний фиксатор, с помощью которых они двигаются, как дюймовые черви . [20]
- Заказать Hoplonemertea. Насчитывает 650 видов. Они живут в придонных и пелагических морских водах, в пресных водах и на суше. Они питаются комменсализмом и паразитизмом и вооружены стилетом (ами) [20]
- Подотряд Monostilifera. Включает 500 видов с одним центральным стилетом . Некоторые используют стилет для передвижения, а также для захвата добычи. [20]
- Подотряд Polystilifera. Включает около 100 пелагических и 50 бентосных видов. Их подушечки несут множество крошечных стилетов. [20]
История эволюции [ править ]
Летопись окаменелостей [ править ]
Поскольку немертины в основном имеют мягкое тело, можно ожидать, что их окаменелости будут крайне редкими. [10] [40] Knaust (2010) сообщили о немертины окаменелости и следы от среднего триаса в Германии . [41] Можно было ожидать, что стилет немертины окаменел, поскольку он состоит из минерального фосфата кальция , но никаких окаменелых стилетов обнаружено не было. [10] [40]
Среднего кембрия ископаемых Amiskwia из Burgess Shale был классифицирован как немертины, на основе сходства некоторых необычных бассейнов немертин глубоководных, но лишь немногие палеонтологи принять эту классификацию как окаменелости Burgess Shale не показывают никаких признаков rhynchocoel , ни кишечных отростков. [40] [42]
Knaust & Desrochers (2019) сообщили об окаменелостях червеобразных организмов с широким диапазоном морфологии, встречающихся на плоскостях напластования, из позднеордовикской ( катианской ) формации Вореаль ( Канада ). У особей, сохраняющих передний конец тела, этот конец заострен или закруглен, с ринхоцелем с хоботком, что характерно для немертинов. Авторы приписали эти окаменелости немертидам и интерпретировали их как самую старую находку группы, о которой сообщалось до сих пор. Тем не менее, Knaust & Desrochers предупредили, что частично сохранившиеся предполагаемые окаменелости немертинов могут в конечном итоге оказаться окаменелостями турбеллярий или кольчатых червей.. [43]
Было высказано предположение, что Archisymplectes , одно из животных пенсильванского возраста из Мазон-Крик в северном и центральном Иллинойсе , может быть немертином. [44] Эта окаменелость, однако, сохраняет только очертания «червя», [40] и нет никаких доказательств наличия хоботка, [45] поэтому нет уверенности в том, что это представляет собой немертиец. [40]
Внутри Немертя [ править ]
| Немертя |
| ||||||||||||||||||||||||
выделяет «Anopla», которые являются парафилетическими . [46]
Нет сомнений в том, что тип Nemertea является монофилетическим (это означает, что тип включает всех и только потомков одного предка, который также был членом этого типа). В синапоморфии (Trait разделяют предка и всех его потомков, но не другими группами) включают выворачивающемуся хобот , расположенные в rhynchocoel. [47]
В то время как Рупперт, Фокс и Барнс (2004) рассматривают Palaeonemertea как монофилетические [46], Толлессон и Норенбург (2003) рассматривают их как парафилетические и базальные (содержат предков более поздних клад). [47] невооружённые немертин ( «без оружия») представляют собой эволюционный класс из немертин без стилетов (содержащего Heteronemertea и Palaeonemerteans), в то время как вооружённые немертин ( «вооруженного») являются монофилетическими, но найти , что палеонемертин вдвойне парафилетические, дав начало и Heteronemertea и Enopla. [46] [47] Рупперт, Фокс и Барнс (2004) рассматривают Bdellonemertea как кладуотдельно от Hoplonemertea , [46] в то время как Thollesson и Norenburg (2003) полагают, что Bdellonemertea являются частью Monostilifera (с одним активным стилетом), которые находятся в пределах Hoplonemertea, что подразумевает, что «Enopla» и «Hoplonemertea» являются синонимами для та же ветка дерева. [47] Polystilifera (с множеством крошечных стилетов) монофилетичны. [46] [47]
Отношения с другими типами [ править ]
В англоязычных письмах немертины традиционно рассматриваются как акоеломатные билатерии, которые наиболее тесно связаны с плоскими червями (Platyhelminthes). Эти пред- кладистика анализа , как подчеркнута общие признаки: multiciliated (с несколькими ресничками на клетку), железистый эпидермисом; палочковидные секреторные тела или рабдиты; лобные железы или органы; протонефридии ; и организационная структура органа управления. [48] Тем не менее, многокомпонентные эпидермальные клетки и клетки эпидермальных желез также обнаруживаются у Ctenophora , Echiura , Sipuncula , Annelida , Mollusca и других таксонов.. Рабдиты nemertea имеют структуру, отличную от таковой у плоских червей в микроскопическом масштабе. Лобные железы или органы плоских червей сильно различаются по строению, и подобные структуры появляются у мелких морских кольчатых червей и личинок энтопрокта . Протонефридии немертеи и плоских червей различаются по строению, [48] и положению - пламенные клетки немертеи обычно находятся в стенках сосудов с жидкостью и обслуживаются «стоками», из которых отходы выходят по небольшому количеству трубок. через кожу [24], в то время как клетки пламени плоских червей разбросаны по всему телу. [49] Строгие сравнения не показывают синапоморфий немертин и платигельминтов нефридий. [48]
В соответствии с более поздними анализами, в развитии немертин эмбрионов, экта мезодерма (внешняя часть мезодермы, который является слоем , в котором большинство внутренних органов построены) получают из клеток , меченные 3a и 3b, и эндомезодерм (внутренняя часть мезодермы) происходит из 4d клетки. Некоторая часть эктомезодермы кольчатых червей , эхиуран и моллюсков происходит из клеток 3a и 3b, тогда как эктомезодерма поликладных плоских червей происходит из клетки 2b, а плоские черви acoel не продуцируют эктомезодерму. У немертинов пространство между эпидермисом и кишечником в основном заполнено хорошо развитыми мышцами, встроенными в неклеточную соединительную ткань.. Эта структура похожа на структуру, обнаруженную у более крупных плоских червей, таких как поликлады и триклады , но аналогичная структура мышц стенки тела, встроенная в неклеточную соединительную ткань, широко распространена среди Spiralia (животных, у которых ранние клеточные деления образуют спиральный узор), таких как сипункулид , echiurans и многие аннелиды. [48]
| Bilateria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Родство Nemerteans с Annelida (включая Echiura, Pogonophora , Vestimentifera и, возможно, Sipuncula) и Mollusca делает ленточных червей членами Lophotrochozoa , которые включают примерно половину существующих типов животных. [52] Группы Lophotrochozoa: те животные, которые питаются с использованием лофофоров ( Brachiopoda , Bryozoa , Phoronida , Entoprocta ); филы, в которых эмбрионы большинства членов развиваются в личинки трохофор (например, Annelida и Mollusca); и некоторые другие типы (такие как Platyhelminthes, Sipuncula, Gastrotricha, Gnathostomulida, Micrognathozoa, Nemertea, Phoronida, Platyhelminthes и Rotifera ). [50] [52] Эти группировки основаны на молекулярной филогении , которая сравнивает участки ДНК и РНК организмов . В то время как анализ молекулярной филогении уверен, что члены Lophotrochozoa более тесно связаны друг с другом, чем с нечленами, отношения между членами в основном неясны. [50] [52]
Большинство первичноротых фил вне спиральных члены Ecdysozoa ( «животные , которые линяют »), которые включают в себя членистоногие , нематода и приапулиду . Большинство других двухсторонних типов находятся в Deuterostomia , включая иглокожие и хордовые . Acoelomorpha , которые не являются ни первичноротыми , ни вторичноротых, рассматривается как базальные билатерии. [50] [52] [53]
См. Также [ править ]
- Emplectonema neesii
- Lineus longissimus
- Parborlasia corrugatus
Примечания [ править ]
- ^ Рупперт, Фокс и Барнс относятся к Lineus longissimus длиной 54 метра (177 футов), выброшенной на берег после шторма у Сент-Эндрюса в Шотландии. [11] По другим оценкам, около 30 метров (98 футов). [12] Зоологам крайне сложно измерить этот вид. [13] Для сравнения:
- Самый длинный зарегистрированный синий кит составил 33,58 метра (110,2 фута). [14]
- Эти динозавры Argentinosaurus и Patagotitan оцениваются примерно 35 метров (115 футов) и 31 метров (102 футов) , соответственно. [15]
- Экземпляр арктической гигантской медузы Cyanea capillata arctica имел длину 36,5 метра (120 футов). [16]
- ^ Sipuncula были объединены в Annelida в 2007 году. [51]
Ссылки [ править ]
- ^ a b c Скотт, Томас (1996). «Немертини, Ринхоцела, Немертея, Немертинея» . Краткая энциклопедия биологии . Вальтер де Грюйтер. С. 815–816 . ISBN 978-3-11-010661-9.
- ^ "Немертя" . Интегрированная система таксономической информации . Проверено 18 февраля 2011 года .
- ^ Седхаген, Томас; Пер Сундберг (1986). "Ранее неизвестное сообщение о немертине в литературе". Архивы естествознания . 13 : 7–8. DOI : 10.3366 / anh.1986.13.1.7 . ISSN 0260-9541 .
- ^ а б Р. Гибсон (1995). «Немертинские роды и виды мира: аннотированный контрольный список исходных названий и цитат из описаний, синонимов, текущего таксономического статуса, местообитаний и зарегистрированного зоогеографического распространения». Журнал естественной истории . 29 (2): 271–561. DOI : 10.1080 / 00222939500770161 .
- ^ Барнс, Ричард Стивен Кент (2001). «Черви» . Беспозвоночные: синтез . Вили-Блэквелл. С. 81–83. ISBN 978-0-632-04761-1. Проверено 27 января 2011 .
- ^ Paps J, J Baguñà, Riutort M (апрель 2009). «Внутренняя филогения Lophotrochozoa: новые идеи из современного анализа ядерных рибосомных генов» . Труды Королевского общества B . 276 (1660): 1245–54. DOI : 10.1098 / rspb.2008.1574 . PMC 2660960 . PMID 19129141 .
- Перейти ↑ Jimenez-Guri E, Paps J, Garcia-Fernandez J, Salo E (2006). «Гены Hox и ParaHox у Nemertodermatida, основной двуногой клады» . Международный журнал биологии развития . 50 (8): 675–9. DOI : 10,1387 / ijdb.062167ej . PMID 17051477 .
- ^ Б с д е е г Мур, Джанет; Оверхилл, Райт (2006). «Глава 7 - Немертя» . В Raith Overhill (ред.). Введение в беспозвоночных (2-е изд.). Издательство Кембриджского университета. С. 75–84. ISBN 978-0-521-85736-9. Проверено 31 января 2011 года .
- ^ a b c d e f g h i j k l m n o p q r s t Ruppert, EE, Fox, RS, and Barnes, RD (2004). «Немертя». Зоология беспозвоночных (7-е изд.). Брукс / Коул. С. 271–274 . ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов ( ссылка )
- ^ Б с д е е г ч я J K L Уокер, JC; Андерсон, Д.Т. (1998). «Platyhelminthes, Nemertea, Entoprocta и Gnathostomulida». В Д. Т. Андерсон (ред.). Зоология беспозвоночных (1-е изд.). Издательство Оксфордского университета, Австралия. С. 79–85. ISBN 978-0-19-553941-7.
- ^ Carwardine, Марк (1995). Книга рекордов Гиннеса . Издательство Guinness. п. 232. ISBN. 978-0-85112-658-6.
- ^ а б Гибсон, Рэй. «Тип Nemertea (Nemertinea, Nemertini, Rhynchocoela)» . Вудс-Хоул , Массачусетс : Морская биологическая лаборатория. Архивировано из оригинала на 2011-09-27 . Проверено 30 марта 2011 года .
- ^ "Король червей" . Энциклопедия водного мира: морская звезда . Корпорация Маршалла Кавендиша. 2004. с. 1420. ISBN 978-0-7614-7418-0. Проверено 30 марта 2011 года .
- ^ Симмондс, Марк (2007). Киты и дельфины мира . Издательство New Holland. п. 155. ISBN 978-1-84537-820-2. Проверено 27 января 2011 .
- ^ Пол, Грегори С. (2019). «Определение самого крупного известного наземного животного: критическое сравнение различных методов восстановления объема и массы вымерших животных» (PDF) . Летопись музея Карнеги . 85 (4): 335–358. DOI : 10.2992 / 007.085.0403 . S2CID 210840060 .
- ^ Carwardine, Марк (2008). Записи о животных . Издательство Стерлинг. п. 241. ISBN. 978-1-4027-5623-8. Проверено 27 января 2011 .
- ^ Хадерли, Юджин Клинтон (1980). Роберт Х. Моррис (ред.). Приливные беспозвоночные Калифорнии . Издательство Стэнфордского университета. С. 85–90. ISBN 978-0-8047-1045-9. Проверено 26 января 2011 .
- ^ Мартин, Гэри Г. (1978). «Новая функция рабдитов: выработка слизи для скольжения ресничек». Зооморфология . 91 (3): 235–248. DOI : 10.1007 / BF00999813 . S2CID 206787592 .
- ^ Б с д е е Ruppert, EE, Fox, RS, и Barnes, RD (2004). «Немертя». Зоология беспозвоночных (7-е изд.). Брукс / Коул. С. 274–275 . ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов ( ссылка )
- ^ a b c d e f g h i j k l Ruppert, EE, Fox, RS, and Barnes, RD (2004). «Немертя». Зоология беспозвоночных (7-е изд.). Брукс / Коул. п. 279 . ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов ( ссылка )
- ^ a b c d e е McDermott, J .; Памела Роу (1985). «Питание, пищевое поведение и кормовая экология немертинов» . Американский зоолог . 25 (1): 113–125. DOI : 10.1093 / ICB / 25.1.113 .
- ^ Роу, Памела; Норенбург, Джон Л .; Маслакова, Светлана (2007). Sol Felty Light; Джеймс Т. Карлтон (ред.). Руководство Лайт и Смита: Приливные беспозвоночные от Центральной Калифорнии до Орегона . Калифорнийский университет Press. С. 221–233. ISBN 978-0-520-23939-5. Проверено 4 февраля 2011 года .
- ^ Свет, Сол Felty (1974). «Тип Nemertea (Rhynchocoela)» . Приливные беспозвоночные побережья Центральной Калифорнии (2-е изд.). Калифорнийский университет Press. С. 55–58. ISBN 978-0-520-00750-5. Проверено 22 февраля 2011 года .
- ^ Б с д е е г Ruppert, EE, Fox, RS, и Barnes, RD (2004). «Немертя». Зоология беспозвоночных (7-е изд.). Брукс / Коул. С. 275–276 . ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов ( ссылка )
- ^ a b c d e Мур, Джанет; Гибсон, Рэй (2001). Немертя . Энциклопедия наук о жизни . Джон Вили и сын. DOI : 10.1038 / npg.els.0001586 . ISBN 978-0470016176.
- ^ Перес-Помарес, Хосе М .; Хуан М. Гонсалес-Роса; Рамон Муньос-Чапули (2009). «Построение сердца позвоночных - эволюционный подход к развитию сердца» . Международный журнал биологии развития . 53 (8–9–10): 1427–1443 [1430]. DOI : 10.1387 / ijdb.072409jp . PMID 19247975 . Проверено 29 мая 2011 года . [ постоянная мертвая ссылка ]
- Перейти ↑ Anderson, DT (1998). «Тип беспозвоночных». В Д. Т. Андерсон (ред.). Зоология беспозвоночных (1-е изд.). Издательство Оксфордского университета, Австралия. п. 4. ISBN 978-0-19-553941-7.
- ^ Ruppert, Е. Е., Fox, RS, и Барнс, RD (2004). «Введение в Bilateria». Зоология беспозвоночных (7-е изд.). Брукс / Коул. С. 212–214 . ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов ( ссылка )
- ^ Б с д е е Ruppert, EE, Fox, RS, и Barnes, RD (2004). «Немертя». Зоология беспозвоночных (7-е изд.). Брукс / Коул. п. 276 . ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов ( ссылка )
- ^ a b c d Маслакова, Светлана А. (июль 2010 г.). «Изобретение личинки пилидиума в совершенно хорошем спиральном типе Nemertea» . Интегративная и сравнительная биология . 50 (5): 734–743. DOI : 10.1093 / ICB / icq096 . PMID 21558236 .
- ^ a b c Барнс, Роберт Д. (1982). Зоология беспозвоночных . Филадельфия, Пенсильвания: Holt-Saunders International. С. 252–262. ISBN 978-0-03-056747-6.
- ^ Рассел, Питер Дж .; Вулф, Стивен Л .; Hertz, Paul E .; Сеси Старр (2008). «Фоторецепторы и зрение» . Биология: динамическая наука . 3 . Cengage Learning. С. 894–895. ISBN 978-0-495-01034-0. Проверено 31 января 2011 года .
- ^ Рассел, Питер Дж .; Вулф, Стивен Л .; Hertz, Paul E .; Сеси Старр (2008). «Фоторецепторы и зрение» . Биология: динамическая наука . 3 . Cengage Learning. п. 889. ISBN. 978-0-495-01034-0. Проверено 31 января 2011 года .
- ^ Shinobe, Shotaro; Учида, Шота; Мори, Хидеаки; Окочи, Исаму; Чиба, Сатоши (2017). «Ухудшение почвенных ракообразных в объекте всемирного наследия, вызванное землей немертин» . Научные отчеты . 7 (1): 12400. DOI : 10.1038 / s41598-017-12653-4 . PMC 5622052 . PMID 28963523 .
- ^ a b c d e f g h i Рупперт, EE, Fox, RS, и Barnes, RD (2004). «Немертя». Зоология беспозвоночных (7-е изд.). Брукс / Коул. С. 276–278 . ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов ( ссылка )
- ^ «Гонодукт - Медицинское определение» . Merriam-Webster, Incorporated . Проверено 31 января 2011 года .
- ^ Hiebert, Laurel S .; Маслакова, Светлана Александровна (11 апреля 2015 г.). «Hox-гены образуют передне-заднюю ось молоди, но не личинки у максимально непрямого развивающегося беспозвоночного, Micrura alaskensis (Nemertea)» . BMC Biology . 13 (1): 23. DOI : 10,1186 / s12915-015-0133-5 . PMC 4426647 . PMID 25888821 .
- ^ Ховарт, Фрэнсис G .; Джанет Мур (1983). «Земля nemertine Argonemertes dendyi (Dakin) на Гавайях (Nemertinea: Hoplonemertinea: Prosorhochmidae)» (PDF) . Тихоокеанская наука . 37 (2): 141–144. Архивировано из оригинального (PDF) 4 ноября 2018 года . Проверено 17 февраля 2011 .
- ^ Шинобе, S; Учида, S; Мори, Н; Окочи, I; Чиба, S (2017). «Ухудшение почвенных ракообразных в объекте всемирного наследия, вызванное землей немертин» . Научные отчеты . 7 (1): 12400. DOI : 10.1038 / s41598-017-12653-4 . PMC 5622052 . PMID 28963523 .
- ^ Б с д е ф Бен Ваггонером; Аллен Дж. Коллинз (13 июня 2001 г.). «Введение в Немертини» . Калифорнийский университет в Беркли . Архивировано из оригинального 10 декабря 2010 года . Дата обращения 3 февраля 2011 .
- ^ Дирк Кнауст (2010). «Замечательно сохранившиеся бентосные организмы и их следы из грязевой равнины среднего триаса (Muschelkalk)». Летая . 43 (3): 344–356. DOI : 10.1111 / j.1502-3931.2009.00196.x .
- ^ Дзик, Ежи (1999). «Эволюционное происхождение асимметрии у ранних многоклеточных животных» . В Дьюла Пали; Лучано Калиоти; Клаудиа Зукки (ред.). Достижения в области биохиральности . Эльзевир. п. 165. ISBN 978-0-08-043404-9. Проверено 11 февраля 2011 года .
- ^ Dirk Knaust; Андре Дерошер (2019). «Исключительно сохранившийся комплекс мягкотелых пород в карбонатах ордовика острова Антикости, восточная Канада». Гондванские исследования . 71 : 117–128. DOI : 10.1016 / j.gr.2019.01.016 .
- ^ Шрам, Фредерик Р. (сентябрь 1973). «Псевдоеломаты и немертин из штата Иллинойс, Пенсильвания». Журнал палеонтологии . 47 (5): 985–989. JSTOR 1303083 .
- ^ Нильсен, Клаус (2001). «Тип Немертини» . Эволюция животных: взаимосвязи живых типов (2-е изд.). Издательство Оксфордского университета. С. 281–282. ISBN 978-0-19-850681-2. Проверено 11 февраля 2011 года .
- ^ Б с д е е Ruppert, EE, Fox, RS, и Barnes, RD (2004). «Немертя». Зоология беспозвоночных (7-е изд.). Брукс / Коул. С. 279–280 . ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов ( ссылка )
- ^ a b c d e Толлессон, Микаэль; Джон Л. Норенбург (февраль 2003 г.). «Отношения ленточного червя: филогения типа Nemertea» . Труды Королевского общества B . 270 (1513): 407–415. DOI : 10.1098 / rspb.2002.2254 . PMC 1691252 . PMID 12639321 .
- ^ а б в г Turbeville, JM (2002). «Прогресс в биологии немертины: развитие и филогения» . Интегративная и сравнительная биология . 42 (3): 692–703. DOI : 10.1093 / ICB / 42.3.692 . ISSN 1540-7063 . PMID 21708766 . Проверено 29 мая 2011 года .
- ^ Рупперт, EE; Fox, RS; Барнс, RD (2004). Зоология беспозвоночных (7-е изд.). Брукс / Коул. п. 239 . ISBN 978-0-03-025982-1.
- ^ a b c d Галаныч, К. М. (декабрь 2004 г.). «Новый взгляд на филогению животных» (PDF) . Ежегодный обзор экологии, эволюции и систематики . 35 : 229–256. DOI : 10.1146 / annurev.ecolsys.35.112202.130124 .
- ^ Ударил, TH; Schult, N .; Кусен, Т .; Hickman, E .; Блейдорн. C .; McHugh, D .; Галаныч, К.М. (2007). «Филогения аннелид и статус Sipuncula и Echiura» . BMC Evolutionary Biology . 7 (57): 57. DOI : 10.1186 / 1471-2148-7-57 . PMC 1855331 . PMID 17411434 .
- ^ a b c d Гирибет, Гонсало (апрель 2008 г.). «Сборка лофотрохозойского (спиралевидного) древа жизни» . Труды Королевского общества B . 363 (1496): 1513–1522. DOI : 10.1098 / rstb.2007.2241 . PMC 2614230 . PMID 18192183 .
- ^ Кэннон, JT; Веллутини, Британская Колумбия; Smith, J .; Ронквист, Ф .; Jondelius, U .; Хейнол, А. (4 февраля 2016 г.). «Xenacoelomorpha - родственная группа Nephrozoa» . Природа . 530 (7588): 89–93. DOI : 10,1038 / природа16520 . PMID 26842059 . S2CID 205247296 .
Внешние ссылки [ править ]
| В Wikisource есть текст статьи Nemertina из Британской энциклопедии 1911 года . |
- Морская биологическая лаборатория: Тип Nemertea (Nemertinea, Nemertini, Rhynchocoela)
- Немертя LifeDesk
- Видео Nemertea в Пьюджет-Саунд