Похоже, что один из основных авторов этой статьи имеет тесную связь с ее предметом. ( Апрель 2017 г. ) ( Узнайте, как и когда удалить этот шаблон сообщения ) |
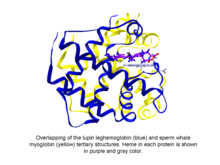
Phytoglobins шаровидных растений (водоросль и земля растение) белки , классифицированные в глобина суперсемействе, которые содержат гем, I . е . протопорфирин IX -Fe, простетическая группа. Самыми простыми известными фитоглобинами являются леггемоглобины , открытые в 1939 году Кубо после спектроскопического и химического анализа красного пигмента клубеньков корня сои . [1] Через несколько десятилетий после доклада Кубо кристаллизация фитоглобина люпина (известного как леггемоглобин) Вайнштейном и его сотрудниками показала, что третичная структура этого белка и миоглобина кашалотабыл удивительно похож, что указывало на то, что фитоглобин, открытый Кубо, действительно соответствовал глобину. [2]
Одной из важных функций фитоглобина является его активность диоксигеназы оксида азота . [3]
Распространение и классификация [ править ]
Фитоглобины (сокращенно Phytogbs) повсеместно распространены в растениях, поскольку они были идентифицированы в водорослях и наземных растениях, включая примитивные мохообразные и эволюционировавшие однодольные и двудольные растения . Их можно классифицировать следующим образом: [3] [4]
- 2/2 Фитоглобины (семейство усеченных глобинов, подсемейство TrHb2)
- Phytogb3: содержится в водорослях и наземных растениях.
- 3/3 Фитоглобины (миоглобиноподобное семейство)
- Phytogb0: базальный тип, обнаруженный в водорослях, мохообразных и голосеменных . [4]
- Phytogb1, 2: встречается только у покрытосеменных .
Не бобовые симбиотические глобины (SymPhytogbs) [4] разбросаны среди Phytogb1 и Phytogb2. [3] Симбиотические глобины обычно снабжают кислородом симбиотические бактерии, которые связывают азот. У бобовых бактерий являются ризобиями , но у некоторых актиноризных растений эту работу выполняет актиномицет Frankia . [3]
Структурные характеристики [ править ]
Фитогены кодируются генами, прерываемыми 3 интронами (хотя у мха Physcomitrella patens был обнаружен содержащий 4 интрона гена phytogb ). [5] Первый и третий интрон генов phytogb локализованы в том же положении, что и гены миоглобина , что предполагает, что гены phytogb и миоглобина произошли от общего предка более 600 миллионов лет назад. [6] Существование второго интрона в генах phytogb было предсказано Go с использованием теоретического анализа [7], который был дополнительно подтвержден клонированием и секвенированием гены lb сои , Маркер и соавторы. [8]
Фитогб представляет собой мономерные белки с молекулярной массой от ~ 17 до ~ 19 кДа. Однако при высоких (более 1 мМ) концентрациях Phytogbs могут образовывать димеры. Полипептидная цепь Phytogbs складывается в определенное расположение из 6-7 спиралей (обозначенных буквами от A до H), известное как глобиновая складка, которая образует гидрофобный карман, в котором расположен гем. Два типа глобиновой складки были идентифицированы в Phytogbs: 3/3- и 2/2-складчатость, [9] [10], где спирали A, E и F перекрываются со спиралями B, G и H и спиралями B и E. перекрываются со спиралями G и H соответственно.
Как и другие глобины, гем-Fe в Phytogbs координируется в проксимальной области с помощью аминокислоты His (называемой проксимальной His). Дистальная область гем-Fe может быть занята множеством лигандов (таких как кислород и оксид азота ) или дистальной (часто His) аминокислотой, уступая место пента- или гексакоординированным Phytogbs, соответственно. Phytogbs2, SymPhytogbs и Lbs являются преимущественно пента-координатами, тогда как Phytogbs1 преимущественно гексакоординатными, а Phytogbs0 и Phytogbs3 представляют собой комбинацию пента- и гексакоординат. [4] Координация гем-Fe важна для функции Phytogb (и других глобинов), потому что она регулирует скорость связывания и высвобождения лиганда как следствие кинетических констант k on иk выкл соответственно. Например, сродство Lb сои и Phytogb1 риса к O 2 ( K O 2 ) является умеренным и очень высоким, потому что k on составляет 130 и 68 мМ -1 с -1 , k off составляет 5,6 и 0,038 с -1, а K O 2 ( я . е . уплотнительное 2 -affinity в результате к на / K выключен ) составляет 23 и 1800 мМ -1 , соответственно. [11] Это указывает на то, что Lb сои может функционировать как O2- хранение или -транспортный белок, и что функция Phytogb1 риса (и других гексакоординированных Phytogbs) может отличаться от транспорта O 2, потому что высокое сродство этого белка к O 2 является результатом чрезвычайно низкой константы k off . [12]
Синтез и постулируемые функции [ править ]
Как и другие глобины, пентакоординированные фитогбы обратимо связываются и транспортируют О 2 . Функция Lbs в узелках была выяснена в 1974 году Виттенбергом, Эпплби и другими. [13] В клубеньках концентрация Lbs очень высока, поскольку они соответствуют ~ 30% от общего количества растворимых белков. Очевидная функция Lbs в клубеньках заключается в облегчении диффузии O 2 к дышащим бактероидам для фиксации азота . В то же время Lb способствует поддержанию низких уровней O 2 (~ 10 нМ), чтобы избежать инактивации чувствительной к O 2 нитрогеназы, которая фиксирует атмосферный азот. [14]
Кроме того, Phytogbs связывают другие газообразные лиганды, в первую очередь оксид азота (NO), и проявляют активность NO- диоксигеназы . [15] Работа Хилла и его сотрудников за последние 15 лет показала, что уровни эндогенного NO варьируются в зависимости от концентрации Phytogbs1 в трансгенной кукурузе и люцерне. [16]Основываясь на этих наблюдениях, эти авторы предположили, что функция насыщенных кислородом Phytogbs заключается в модуляции уровней NO посредством активности NO-диоксигеназы и в косвенной регуляции широкого спектра функций клетки, которые модулируются уровнями NO. Кислородсодержащие фитоглобины класса 1, реагирующие с NO с образованием нитрата, представляют собой основной механизм поглощения NO растениями. Цикл, включающий нитратредуктазу, восстановление нитрита до NO, поглощение NO фитоглобином, был определен как цикл фитоглобин-NO . [17] Его работа приводит к поддержанию окислительно-восстановительного и энергетического статуса во время гипоксии и приводит к снижению производства этанола и молочной кислоты. [18]
Phytogbs0, 1, 2 и 3 синтезируются в очень низких концентрациях в различных (эмбриональных и вегетативных) органах растений. [19] [20] [21] Однако концентрация Phytogbs увеличивается в растениях, подвергающихся определенным стрессовым условиям, таким как затопление [22] и ограничение света. [23] Следовательно, некоторые Phytogbs считаются чувствительными к стрессу белками растений.
Эволюция [ править ]
Части этой статьи (те, которые связаны с филогенезом Беканы, отвергают общее происхождение Lbs и SymPhytogbs в Phytogb1), нуждаются в обновлении . ( Ноябрь 2020 г. ) |
Работа Арредондо-Питера, Виноградова и его сотрудников прояснила основные события, которые могли произойти во время эволюции Phytogbs. [24] [25] [26] Несмотря на то, Phytogbs произошли от общего предка , предшествующего происхождения эукариот ( я . Е . В эубактерийдомен), в растениях были идентифицированы две эволюционные линии Phytogbs: линия (3 / 3-) Phytogb0, 1 и 2 и линия SymPhytogb и Lb и линия (2 / 2-) Phytogb3. По-видимому, Phytogbs0 были предками Phytogbs1 и 2, а SymPhytogbs и Lbs произошли от предка Phytogb1 как приспособление к симбиотической азотфиксации. Сравнительный анализ мха Phytogb0 со структурой Phytogb1 риса и Lb сои показал, что основные эволюционные изменения, которые, вероятно, произошли во время эволюции Phytogbs0, 1 и 2, SymPhytogbs и Lbs, были (i) переходом гекса-координаты в пента-координату в геме. группы, (ii) уменьшение длины CD-петли и N- и C-концов, и (iii) уплотнение белка в глобулярную структуру. Более того,структура глобинового домена Phytogbs3 очень похожа на структуру бактериальных усеченных глобинов, что указывает на то, что 2/2-фолдинг сохраняется во время эволюции Phytogbs3. Однако глобиновый домен наземного растения Phytogbs3 фланкирован внеаминокислотными последовательностями, которые, вероятно, произошли от предка наземного растения Phytogbs3. Функциональное и эволюционное значение дополнительных аминокислотных последовательностей в структуре Phytogbs3 до сих пор не известно.
Технологические приложения [ править ]
Фитогбы потенциально полезны для ряда биотехнологических применений. Например, вскоре после открытия риса Phytogbs1 предполагалось использовать эти белки в качестве датчиков O 2 в электронных устройствах. Кроме того, сверхэкспрессия Phytogbs была предложена в качестве стратегии повышения устойчивости сельскохозяйственных культур к определенным стрессовым условиям, таким как затопление сельскохозяйственных культур, [27] [28], поскольку это связывает ранний сигнальный этилен наводнения с низкой кислородной реакцией растений. [29] [30] Недавно Phytogbs были рассмотрены кандидатами на разработку кровезаменителей и добавок в овощные гамбургеры .
Заметки [ править ]
Ссылки [ править ]
- ^ Кубо Х., Uber hamoprotein aus den wurzelknollchen von leguminosen, Acta Phytochim. (Токио), 11 (1939) 195-200.
- ^ Вайнштейн Б.К., Арутюнян Э.Х., Куранова И.П., Борисов В.В., Н.И.Сосфенов, Павловский А.Г., Гребенко А.И., Конарева Н.В., Структура леггемоглобина из клубеньков корня люпина при разрешении 5 А., Природа, 254 (1975) 163-164
- ^ a b c d e f Бекана, Мануэль; Юруэла, Инмакулада; Сарат, Гаутама; Каталон, Пилар; Харгроув, Марк С. (сентябрь 2020 г.). «Гемоглобины растений: путь от одноклеточных зеленых водорослей к сосудистым растениям» . Новый фитолог . 227 (6): 1618–1635. DOI : 10.1111 / nph.16444 . PMID 31960995 .
- ^ а б в г Хилл Р., Харгроув М.С., Арредондо-Питер Р. (2016). «Фитоглобин: новая номенклатура глобинов растений, принятая глобиновым сообществом на XVIII конференции 2014 года по связыванию кислорода и чувствительным белкам» . F1000 Исследования . 5 : 212. DOI : 10,12688 / f1000research.8133.1 . PMC 4792203 . PMID 26998237 . CS1 maint: использует параметр авторов ( ссылка )
- ^ Мартинес-Окампо Ф., Васкес-Лимон С., Арредондо-Питер Р., Обнаружение, ПЦР-амплификация и характеристика несимбиотического гена гемоглобина, содержащего 4 интрона, из мха Physcomitrella patens ., Global J. Biochem., 3 (2012) 12
- ^ Hardison RC, Краткая история гемоглобинов: растений, животных, протистов и бактерий., Proc. Natl. Акад. Sci. USA., 93 (1996) 5675-5679
- ^ Го М., Корреляция экзонных областей ДНК со структурными единицами белка в гемоглобине., Nature, 291 (1981) 90-92.
- ^ Hyldig-Nielsen JJ, Jensen E., Paludan K., Wiborg O., Garret R., Joergersen PL, Marker KA, Первичные структуры двух генов леггемоглобина сои, Nucleic Acids Res., 10 (1982) 689-701 .
- ^ Харгрув М., Брукер Е.А., Стец Б., Сарат Г., Arredondo-Питер Р., Klucas Р.В., Olson JS, PhilipsJr. Г. Н., Кристаллическая структура несимбиотического гемоглобина., Структура, 8 (2000) 1005-1014.
- ↑ Reeder BJ, Hough MA, Структура несимбиотического растительного гемоглобина класса 3 из Arabidopsis thaliana обнаруживает новое N-концевое спиральное удлинение., Acta Crystallogr., D70 (2014) 1411-1418.
- ^ Arredondo-Питер Р., Харгрув М.С., Moran JF, Сарат Г., Klucas Р.В., гемоглобинов завод, завод Physiol., 118 (1998) 1121-1126.
- ^ Arredondo-Peter R., Hargrove MS, Sarath G., Moran JF, Lohrman J., Olson JS, Klucas RV, Гемоглобины риса: клонирование гена, анализ и кинетика связывания кислорода рекомбинантного белка, синтезированного в Escherichia coli ., Plant Physiol., 115 (1997) 1259-1266.
- ^ Виттенберг Дж. Б., Бергерсен Ф. Дж., Эпплби К. А., Тернер Г. Л., Облегченная диффузия кислорода: роль леггемоглобина в фиксации азота бактероидами, выделенными из клубеньков корня сои., J. Biol. Chem., 249 (1974) 4057-4066
- ^ Appleby CA, Леггемоглобин идыхание Rhizobium ., Annu. Rev. Plant Physiol., 35 (1984) 443-478.
- ^ Smagghe BJ, Trent JT, Hargrove MS, NO-диоксигеназная активность в гемоглобинах повсеместна in vitro , но ограничена снижением in vivo ., PLoS One, 3 (2008) doi: 10.1371 / journal.pone.0002039.
- ^ Hill RD, Несимбиотические гемоглобины - Что происходит помимо поглощения оксида азота ?, AoB Plants, Pls004 (2012) doi: 10.1093 / aobpla / pls1004
- ^ Игамбердиев, AU; Барон, К .; Manac'h-Little, N .; Стойменова, М .; Хилл, RD (2005). «Цикл гемоглобина / оксида азота: участие в стрессе наводнения и влияние на передачу сигналов гормона» . Летопись ботаники . 96 (4): 557–564. DOI : 10.1093 / Aob / mci210 . ISSN 0305-7364 . PMC 4247025 . PMID 16027133 .
- ^ Игамбердиев, AU; Хилл, RD (2004). «Нитраты, NO и гемоглобин в адаптации растений к гипоксии: альтернатива классическим путям ферментации» . Журнал экспериментальной ботаники . 55 (408): 2473–2482. DOI : 10.1093 / JXB / erh272 . ISSN 0022-0957 . PMID 15448180 .
- ^ Гаррохо-Виллегас В., Арредондо-Питер Р., Молекулярное клонирование и характеристика несимбиотического гемоглобинамха ( Ceratodon purpureus ) дает представление о ранней эволюции несимбиотических гемоглобинов растений., Mol. Биол. Evol., 25 (2008) 1482-1487.
- ^ Росс EJH, Shearman L., Mathiesen M., Zhou J., Arredondo-Peter R., Sarath G., Klucas RV, Несимбиотические гемоглобины синтезируются во время прорастания и в дифференцирующихся типах клеток., Protoplasma, 218 (2001) 125-133.
- ^ Watts RA, Hunt PW, Hvitved AN, Hargrove MS, Peacock WJ, Dennis ES, гемоглобин из растений, гомологичных усеченным гемоглобинам микроорганизмов., Proc. Natl. Акад. Sci. USA., 98 (2001) 10119-10124.
- ^ Taylor ER, Nie XZ, MacGregor AW, Hill RD, Ген гемоглобина злаков экспрессируется в тканях семян и корней в анаэробных условиях, Plant Mol. Биол., 24 (1994) 853-862
- ^ Лира-Руан В., Сарат Г., Клукас Р.В., Арредондо-Питер Р., Синтез гемоглобинов врастенияхриса ( Oryza sativa var. Jackson), растущих в нормальных и стрессовых условиях., Plant Sci., 161 (2001) 279 -287.
- ^ Васкес-Лимон К., Хугевийс Д., Виноградов С.Н., Арредондо-Петер Р., Эволюция гемоглобинов наземных растений, Plant Sci., 191-192 (2012) 71-81.
- ^ Виноградов С.Н., Фернандес И., Хугевийс Д., Арредондо-Петер Р., Филогенетические отношения 3/3 и 2/2 гемоглобинов в геномах Archaeplastida с бактериальными и другими гемоглобинами эукариот, Мол. Завод, 4 (2011) 42-58.
- ^ Виноградов С.Н., Хугевийс Д., Арредондо-Питер Р., Каково происхождение и филогения гемоглобинов растений ?, Commun. Интегрировать. Биол., 4 (2011) 443-445.
- ^ Анджейчак, О.А.; Havelund, JF; Ван, WQ; Ковальчук, С; Хагенсен, CE; Hasler-Sheetal, H; Дженсен, Онтарио; Rogowska-Wrzesinska, A; Møller, IM; Hebelstrup, KH (24 февраля 2020 г.). «Гипоксический протеом и метаболом ячменя ( Hordeum vulgare L.) с праймингом фитоглобином и без него» . Международный журнал молекулярных наук . 21 (4): 1546. DOI : 10,3390 / ijms21041546 . PMC 7073221 . PMID 32102473 .
- ^ Мира, Мохамед М .; Хилл, Роберт Д.; Стасолла, Клаудио (ноябрь 2016 г.). «Фитоглобины улучшают гипоксический рост корней, облегчая гибель клеток апикальной меристемы» . Физиология растений . 172 (3): 2044–2056. DOI : 10.1104 / pp.16.01150 . PMC 5100795 . PMID 27702845 .
- ^ Хартман, Иоганн Герард Вильгельм (2020-01-22). Сигнал раннего наводнения этилен позволяет растениям адаптироваться к стрессу с низким содержанием кислорода . Universiteit Utrecht. hdl : 1874/387763 . ISBN 978-90-393-7231-9.
- ^ Хартман, S; Лю, Z; van Veen, H; Vicente, J; Reinen, E; Martopawiro, S; Чжан, Х; van Dongen, N; Босман, Ф; Bassel, GW; Виссер, EJW; Бейли-Серрес, Дж; Теодулу, Флорида; Hebelstrup, KH; Гиббс, диджей; Холдсворт, MJ; Sasidharan, R; Voesenek, LACJ (5 сентября 2019 г.). «Этилен-опосредованное истощение оксида азота предварительно адаптирует растения к стрессу гипоксии» . Nature Communications . 10 (1): 4020. Bibcode : 2019NatCo..10.4020H . DOI : 10.1038 / s41467-019-12045-4 . PMC 6728379 . PMID 31488841 .