| Одонтогрифус | |
|---|---|
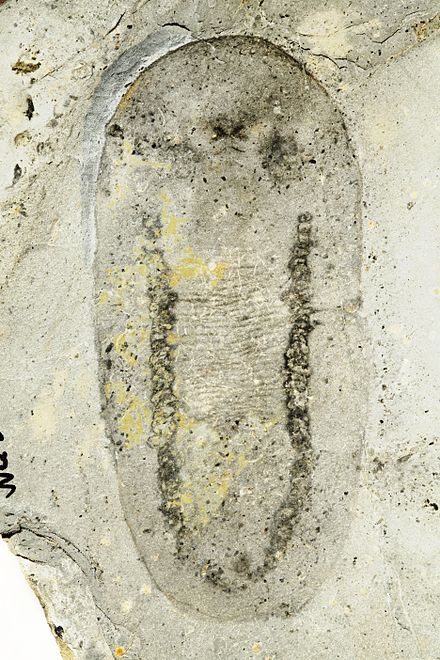 | |
| Одонтогрифус из сланцев Берджесс. От Смита (2014). [2] | |
 | |
| Несколько разложившаяся окаменелость одонтогрифуса с радулой и кишечником. От Смита (2014) [2] | |
| Научная классификация | |
| Королевство: | Animalia |
| Суперфилум: | Лофотрохозоа |
| Семья: | Odontogriphidae Conway Morris, 1976 [1] |
| Род: | Одонтогрифус Конвей Моррис, 1976 [1] |
| Разновидность | |
| |
{kind=link}
{kind=link}
Odontogriphus (буквально «зубчатая загадка») - род мягкотелых животных, известный из среднего кембрия Lagerstätte . Достигая 12,5 сантиметров (4,9 дюйма) в длину, одонтогрифус представляет собой плоское овальное двухстороннее животное, у которого, по-видимому, была единственная мускулистая ступня и «панцирь» на спине, который был умеренно жестким, но из материала, не подходящего для окаменелости .
Первоначально он был известен только по одному экземпляру, но 189 новых находок в годы, непосредственно предшествующие 2006 г., сделали возможным подробное описание. (221 экземпляр Odontogriphus известен из пласта Greater Phyllopod , где они составляют 0,42% сообщества. [3] ) В результате, Odontogriphus занял видное место в дебатах об эволюционном происхождении моллюсков , которые продолжались с 1990 года. кольчатые черви и брахиоподы . Считается, что устройство для кормления Odontogriphus , которое «почти идентично» устройству Wiwaxia , является ранней версией радулы моллюска ,хитиновый «язык», на котором расположены несколько рядов грубых зубов. Следовательно, Odontogriphus и Wiwaxia часто классифицируются как близкородственные истинным моллюскам.
История открытия [ править ]
Чарльз Дулиттл Уолкотт нашел один образец во время одной из своих полевых поездок к сланцу Берджесс между 1910 и 1917 годами. В 1970-х Саймон Конвей Моррис повторно исследовал образец и сделал предварительный вывод о том, что это плавучий лофофорат , другими словами, связанный с предками моллюски , кольчатые черви и брахиоподы . [1] В 2006 г. Caron, Scheltema et al. опубликовал новый анализ, основанный на 189 недавно собранных образцах, все из сланцев Берджесс. [4]
Описание [ править ]
Odontogriphus, по- видимому, был очень редким видом, составляя менее 0,5% отдельных организмов, обнаруженных в одних и тех же ископаемых пластах. Большинство окаменелостей состоит из двух частей расколотого блока породы: верхняя часть представляет собой « слепок » верхней поверхности животного, а нижняя - одну из его нижней стороны. [4]
Одонтогрифус был животным с плоским телом длиной от 3,3 мм (0,13 дюйма) до 125 мм (4,9 дюйма) с параллельными сторонами и полукруглыми концами. Образцы, исследованные Caron, Scheltema et al. (2006) имели одинаковое соотношение длины к ширине независимо от размера. Очертания тела двусторонне симметричны у всех достаточно полных экземпляров, даже у тех, у которых внутренние черты сохранились асимметрично. Caron, Scheltema et al. (2006) интерпретировали это как доказательство того, что у животных на спине были «панцири», которые были достаточно жесткими, чтобы противостоять любым напряжениям, искажающим внутренние особенности, но не были достаточно прочными, чтобы их можно было сохранить путем окаменения - как, например, ногти на пальцах.. Относительно широкие морщины, параллельные друг другу и обычно прямые, у некоторых экземпляров проходят через центральную часть тела. [4]
Caron, Scheltema et al. (2006) нашли доказательства круглого рта на нижней стороне с двумя, а иногда и тремя структурами, несущими зубы, которые они интерпретировали как устройство для кормления и очень похоже на таковое у Wiwaxia . Odontogriphus устройство подачи «s было расположено на средней линии, около 15% от общей длины тела от переднего края ископаемых. [4]Ротовой аппарат состоял из двух-трех рядов, каждый из которых содержал около двух десятков углеродистых зубов, расположенных симметрично относительно медиального зуба, с одним или двумя боковыми зубами, существенно меньшими, чем центральные зубы. Зубы работали, проходя вокруг нижележащего «языка», при этом зубные ряды деформировались. Зубы, вероятно, выкапывали грязь с морского дна, питаясь любым детритом в ней. [5]
По обе стороны от подающего устройства находится круглая конструкция, которую Caron, Scheltema et al. (2006) интерпретируются как слюнные железы . Они также нашли доказательства других частей пищеварительного тракта : пищевода, соединяющегося с задней частью рта; относительно короткий живот ; и прямой, гораздо более длинный кишечник , заканчивающийся анусом в задней части нижней части. По-видимому, большая железа покрывает заднюю часть кишечника. [2] Пара структур по обе стороны кишечника и немного позади желудка, возможно, была гонадой или органами пищеварения. [4]Мышечная ткань, по-видимому, сохранилась в голове и стопе с распределением, поразительно напоминающим то, что наблюдается в современных хитонах . [2]
Окаменелости показали признаки утолщенной центральной структуры, которую Caron, Scheltema et al. (2006) думали, что это нижняя сторона и, вероятно, представляет собой мускулистую подошву, которая была чуть больше половины ширины всего животного. Он имел U-образную форму, с «открытым» концом позади рта и закругленным концом немного впереди заднего края животного. Очевидно, анус был немного впереди закругленного конца. Все края «стопы», кроме переда, были окружены более темными пятнами, которые иногда отделялись от остальной части тела тонким слоем осадка. [4]
Филогения [ править ]
Odontogriphus занял видное место в дебатах об эволюционном происхождении моллюсков , кольчатых червей и брахиопод, которые продолжаются с 1990 года . [6] [7] Карон, Шелтема и др. (2006) интерпретировали устройство питания Odontogriphus как предшественника радулы моллюска.на том основании, что: случайный и менее отчетливый третий зубной ряд выглядел как свидетельство того, что животные вырастали замещающие зубные ряды в задней части рта и сбрасывали изношенные спереди, как это происходит с радулами моллюсков; изолированные пары зубных рядов, которые они обнаружили, не связанные с окаменелостями тела, но в тех же относительных положениях, что и в более полных окаменелостях, предполагают, что они были установлены на довольно жестких поверхностях, таких как хитиновые «пояса» современных радул; они даже обнаружили признаки того, что отброшенные зубные ряды иногда съедались животными. Следовательно, они классифицировали и Odontogriphus , и Wiwaxia с его «почти идентичным» питательным аппаратом, как примитивных родственников моллюсков. [4]
| |||||||||||||||||||||||||||||||||||||||||||
В соответствии с этой классификацией они интерпретировали темные пятна вокруг стопы как жаберные ктенидии , еще одну особенность некоторых моллюсков; и осадок, который иногда появлялся в окаменелостях между ногой и предполагаемыми ктенидиями, предполагал наличие мантийной полости. [4] Они также пришли к выводу, что Odontogriphus был тесно связан с эдиакарским животным Kimberella , [4] чьи окаменелости также показывают признаки довольно жесткой верхней «оболочки», сделанной из материала, который не окаменел , и который был интерпретирован как очень моллюскоподобный организм. [8] [9]
Они продолжили классифицировать халкиерид как почти современных моллюсков, поскольку, по их мнению, «кольчужная» оболочка минерализованных склеритов халькериид была продвижением по сравнению с неминерализованными склеритами Wiwaxia, а также напоминала панцирь некоторых живых аплакофорных моллюсков без панцирей , т.е. Neomeniomorpha . В результате они пришли к выводу, что вся линия моллюсков Kimberella - Odontogriphus - Wiwaxia должна была отличаться от линии кольчатых червей за некоторое время до появления Kimberella в эдиакарский период. [4]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
Это поставило Odontogriphus в центр дебатов, которые продолжались с 1990 года, когда Баттерфилд отрицал, что Wiwaxia была предшественницей моллюсков, и утверждал, что это эволюционная «тетя» кольчатых червей. В частности, он утверждал, что: склериты Wiwaxia внутренне были больше похожи на щетинки многощетинковых кольчатых червей, таких как Canadia, чем на любого предшественника пластин раковин моллюсков; и, по его мнению, устройство питания Wiwaxia было больше похоже на устройство некоторых полихет, чем на радулу моллюска. [11] Карон, Шелтема и др. (2006) считали, что Wiwaxia мало похожа на полихет, поскольку не проявляет признаков сегментации., придатки перед ртом или « ноги » - все это типичные черты полихет. [4]
Несколько месяцев спустя, в 2006 году, Баттерфилд вернулся в бой. Как и в 1990 году, он утверждал, что склериты Wiwaxia внутренне были больше похожи на щетинки многощетинковых кольчатых червей, таких как Canadia, чем на любого предшественника пластин раковин моллюсков; поскольку в статье 2005 г. этот аргумент был преуменьшен комментарием о том, что подобные щетинки также появляются у моллюсков и брахиопод [6], он указал, что модифицированные щетинки покрывают спину только у полихет, и, следовательно , склериты Wiwaxia действительно следует рассматривать. как щетинки полихет. [12]
Кроме того, он утверждал, что выпадение и замена зубных рядов Odontogriphus , сохранение рядов в тех же относительных положениях при изоляции и свидетельство того, что Odontogriphus иногда проглатывал выброшенные зубные ряды, не доказывают, что Odontogriphus был эволюционной «теткой» моллюсков , поскольку эуницидные полихеты также линяют и заменяют свой питательный аппарат (который иногда напоминает радулу [13] ), а иногда и поедают выброшенный материал. Он также сомневался, что два ряда зубов Odontogriphus и Wiwaxiaмог выполнять все функции многорядной радулы - вычищать, улавливать очищенную пищу, сортировать ее и транспортировать к пищеводу. По его мнению, различия между более узким первым зубным рядом и немного более широким вторым как у Odontogriphus, так и у Wiwaxia были не похожи на различия у моллюсковой радулы, у которых гораздо более многочисленные зубные ряды идентичны; вместо этого он утверждал, что эти два ряда напоминают постоянную нижнюю челюсть и плавную верхнюю челюсть современных дорвиллидных полихет. [12]
Хотя Баттерфилд согласился с тем, что темные пятна вокруг стопы служат жабрами , он отрицал, что они были похожи по структуре и способу развития на ктенидии моллюсков. По его мнению, уплощенные останки Odontogriphus образованы относительно жесткими внеклеточными выделениями , такими как челюсти, щетина и жесткая кожа, и не включают чисто или преимущественно клеточные ткани, такие как мышцы или гонады . Поэтому он считал, что органы дыхания по краю стопы Odontogriphus не могут быть ктенидиями моллюсков, поскольку они покрыты чисто клеточной тканью. Вместо этого он предложил , чтобы они могли быть брахиоподовым lophophores, которые представляют собой органы питания, содержащие много внеклеточного материала, или полихетные жабры, которые представляют собой органы дыхания, состоящие в основном из неклеточной кутикулы - оба этих типа структур были обнаружены в сланцах Берджесс , в которых находятся все известные образцы Odontogriphus были обнаружены. [12]
Карон, Шелтема и др. (2006) предположили, что морщины на верхней поверхности образцов Odontogriphus были вызваны волнообразными сокращениями мускульной стопы, напоминающей моллюск. [4] Баттерфилд оспорил это на том основании, что: стопа моллюска также в основном состоит из клеточного материала, который, по его мнению, вряд ли окаменел бы в условиях сланцевого месторождения Берджесс; морщины были слишком прямыми и слишком точно пересекали тело животных; промежутки между ними были такого же размера, как и промежутки между жаброобразными структурами вокруг ступни. Вместо этого он утверждал, что это свидетельство видимой снаружи сегментации, которая встречается у полихет, но не у моллюсков. Он пришел к выводу, что Wiwaxia была эволюционной «теткой» полихет, в то время какОдонтогрифус мог быть эволюционной «теткой» полихет, моллюсков или брахиопод - или даже «двоюродной теткой» всех трех, поскольку он мог быть одним из первых представителей лофотрохозоа , « суперфила », включающего полихет. , моллюски и брахиоподы. [12]
В январе 2007 г. Caron, Scheltema, et al. опубликовали энергичный ответ на аргументы Баттерфилда - ближе к концу они написали: «Многих заблуждений Баттерфилда можно было бы избежать, если бы он воспользовался возможностью изучить весь новый материал, который лег в основу нашего исследования…» Они сказали, что нашли в окаменелостях тела Odontogriphus видны следы перепонки, на которой располагались его зубные ряды; по их мнению, это было явным доказательством базового ленточного узла радулы с равномерно расположенными зубными рядами, что является уникальной особенностью моллюсков. С другой стороны, писали они, челюсти эуницидных полихет лишь отдаленно похожи на радулы, а челюсти других кольчатых челюстей непрерывно растут без замены;и они подтвердили это подробным сравнением Odontogriphus'питающий аппарат с аппаратом дорвиллид полихет, на который, по утверждению Баттерфилда, он похож. В ответ на утверждение Баттерфилда о том, что органы дыхания вокруг стопы не могут быть ктенидиями моллюсков, потому что эти в основном клеточные структуры не окаменели бы в условиях сланцев Берджесс, они написали, что: довольно мягкая клеточная ткань, принадлежащая желудку, окаменела во многих образцах Odontogriphus ; некоторые жабры моллюсков укреплены неклеточным материалом, например, у полиплакофора . Они указали, что морщины, которые появляются на теле при виде сверху, возникают только в средней части, и нет никаких признаков того, что жесткая пластина «панцирь» на спине животного была сегментирована; следовательно, по их мнению, Odontogriphusне мог быть кольчатым червем. С другой стороны, на стопах мертвых хитонов видны морщины . Они утверждали, что Wiwaxia явно не сегментирована, поскольку количество склеритов в трех ее концентрических группах совершенно не совпадает. Они раскритиковали главный аргумент Баттерфилда за « впихивание » Wiwaxia в полихеты, что его склериты секретируются микровиллами; Такие структуры, писали они, также были обнаружены у нескольких групп моллюсков. Наконец, по их мнению, отсутствие «ног» у Wiwaxia исключало близкое родство с полихетами. [7]
В 2008 году Баттерфилд описал набор микро-окаменелости датируются между 515 миллионов лет назад и 510 миллионов лет назад , обнаружили в формировании Mahto в Альберте «s Jasper National Park - это ископаемое кровать 510 М лет старше Burgess Shale в котором единственные известные экземпляры Odontogriphus и Wiwaxiaбыли найдены. Некоторые группы этих окаменелостей показали последовательное расположение, которое он интерпретировал как «сочлененный аппарат» с десятками близко расположенных зубных рядов, очевидно установленных на органической основе, и с заметными признаками износа в рядах на одном конце. Ряды были не совсем идентичны, но он отметил, что некоторые современные моллюски аплакофораны демонстрируют похожие вариации. Он пришел к выводу, что «сочлененный аппарат» - это настоящая радула моллюсков, наиболее похожая на таковых современных аплакофоранов или брюхоногих моллюсков . Затем он прокомментировал контраст между этим устройством и двумя или, реже, тремя широко расположенными и более неоднородными зубными рядами, обнаруженными в окаменелостях Odontogriphus и Wiwaxia , и пришел к выводу, что Odontogriphus иВряд ли Wiwaxia были моллюсками. [14]
Однако исследование 2012 года повторно изучило ротовые органы этих двух родов и выявило проблемы с предыдущими интерпретациями. Что наиболее важно, в ротовой полости есть центральный рахидиевый зуб - ключевая характеристика радулярного аппарата. В свете этой новой реконструкции родство моллюсков кажется хорошо подтвержденным. [5] Дальнейшее подтверждение положения в общей группе моллюсков обеспечивается деталями его мускулатуры и дальнейшими деталями, полученными от его близкого родственника Wiwaxia . [2]
Ссылки [ править ]
- ^ a b c d Конвей Моррис, С. (1976). «Новый кембрийский лофофорат из сланцев Берджесс в Британской Колумбии» (PDF) . Палеонтология . 19 : 199–222. Архивировано из оригинального (PDF) 24 августа 2011 года . Проверено 12 октября 2010 .
- ^ а б в г д Смит, MR (2014). «Онтогенез, морфология и систематика мягкотелого кембрийского моллюска Wiwaxia ». Палеонтология . 57 (1): 215–229. DOI : 10.1111 / pala.12063 .
- ^ Карон, Жан-Бернар; Джексон, Дональд А. (октябрь 2006 г.). «Тафономия сообщества Большого пласта филлопод, сланец Берджесс». ПАЛАИ . 21 (5): 451–65. DOI : 10,2110 / palo.2003.P05-070R . JSTOR 20173022 .
- ^ Б с д е е г ч я J к л м н Caron, JB; Scheltema, A .; Schander, C .; Рудкин, Д. (13.07.2006). «Мягкий моллюск с радулой из среднекембрийских сланцев Берджесс». Природа . 442 (7099): 159–163. Bibcode : 2006Natur.442..159C . DOI : 10,1038 / природа04894 . hdl : 1912/1404 . PMID 16838013 . S2CID 4431853 . Полный предварительный вариант перед публикацией, бесплатно, но без изображений, может быть доступен в Caron, Jean-Bernard; Шельтема, Амели; Шандер, Кристофер; Рудкин, Дэвид (2006). «Мягкий моллюск с радулой из среднекембрийских сланцев Берджесс» (PDF) . Природа . 442 (7099): 159–63. Bibcode : 2006Natur.442..159C . DOI : 10,1038 / природа04894 . hdl : 1912/1404 . PMID 16838013 . S2CID 4431853 . Архивировано из оригинального (PDF) 18 июля 2011 года . Проверено 4 июля 2008 г. .
- ^ а б Смит, MR (2012). "Ротовые части окаменелостей Берджесса сланца Odontogriphus и Wiwaxia : Последствия для предка моллюсков радулы" . Труды Королевского общества B . 279 (1745): 4287–4295. DOI : 10.1098 / rspb.2012.1577 . PMC 3441091 . PMID 22915671 .
- ^ a b Эйбье-Якобсен, Д. (сентябрь 2004 г.). «Переоценка Wiwaxia и полихет из сланца Берджесс». Летая . 37 (3): 317–335. DOI : 10.1080 / 00241160410002027 .
- ^ а б Кэрон, JB; Scheltema, A .; Schander, C .; Рудкин, Д. (январь 2007 г.). «Ответ Баттерфилду о червях стеблевой группы: ископаемые лофотрохозоиды в сланцах Берджесс» . BioEssays . 29 (2): 200–202. DOI : 10.1002 / bies.20527 . PMID 17226817 . S2CID 7838912 . Архивировано из оригинала на 2012-10-13 . Проверено 13 августа 2008 .
- ^ Федонкин, М.А. Вагонер, Б.М. (1997). «Позднедокембрийская окаменелость Kimberella - это моллюскоподобный двухсторонний организм». Природа . 388 (6645): 868–871. Bibcode : 1997Natur.388..868F . DOI : 10.1038 / 42242 . S2CID 4395089 .
- ^ Федонкин, М.А. Simonetta, A .; Иванцов, А.Ю. (2007). «Новые данные о кимберелле , вендском моллюскоподобном организме (Белое море, Россия): палеоэкологические и эволюционные последствия». Геологическое общество, Лондон, специальные публикации . 286 (1): 157–179. Bibcode : 2007GSLSP.286..157F . DOI : 10,1144 / SP286.12 . S2CID 331187 .
- ^ NJ, Баттерфилд (2007-12-18). «Корни и стебли лофотрохозойных». В Budd, GE; Streng, M .; Дейли, AC; Уиллман, С. (ред.). Программа с тезисами . Ежегодное собрание палеонтологической ассоциации . Упсала, Швеция. С. 26–7.
- Перейти ↑ Butterfield, NJ (1990). «Переоценка загадочной окаменелости сланца Берджесс Wiwaxia corrugata (Мэтью) и ее связи с полихетой Canadia spinosa . Уолкотт». Палеобиология . 16 (3): 287–303. DOI : 10.1017 / S0094837300010009 . JSTOR 2400789 .
- ^ а б в г Баттерфилд, штат Нью-Джерси (2006). «Зацепление некоторых« червей »стеблевой группы: ископаемых лофотрохозоидов в сланцах Берджесс» . BioEssays . 28 (12): 1161–6. DOI : 10.1002 / bies.20507 . PMID 17120226 . S2CID 29130876 . Архивировано из оригинала на 2011-08-13 . Проверено 6 августа 2008 .
- ^ Баттфилд указывает на рис. 10F в Цетлине А; Пуршке Г. (2005). «Глотка и кишечник». У Бартоломея Т; Пуршке Г (ред.). Морфология, молекулы и филогения полихет и родственных таксонов (PDF) . С. 199–225. DOI : 10.1007 / s10750-004-1431-Z . S2CID 20398766 . и Рис. 4B в Tzetlin, AB (2005). «Ультраструктурное исследование структур челюстей двух видов Ampharetidae (Annelida: Polychaeta)». Acta Zoologica . 85 (3): 171–180. DOI : 10.1111 / j.0001-7272.2004.00168.x .; ряд челюстей кольчатых червей можно наблюдать в Paxton, H (2009). «Филогения Eunicida (Annelida) на основе морфологии челюстей» (PDF) . Зоосимпозия . 2 : 241–264. DOI : 10.11646 / zoosymposia.2.1.18 . ISSN 1178-9905 .
- ↑ Баттерфилд, Нью-Джерси (май 2008 г.). «Раннекембрийская радула». Журнал палеонтологии . 82 (3): 543–554. DOI : 10.1666 / 07-066.1 . S2CID 86083492 .
Дальнейшее чтение [ править ]
- Бенгтсон, С. (13 июля 2006 г.). «Призрак с укусом». Природа . 442 (7099): 146–147. Bibcode : 2006Natur.442..146B . DOI : 10.1038 / 442146a . PMID 16838007 . S2CID 2579864 .
Внешние ссылки [ править ]
- « Odontogriphus omalus » . Галерея ископаемых сланцевых пород Берджесса . Виртуальный музей Канады. 2011 г.
- Статья в Королевском музее Онтарио об O. omalus
- http://www.cbc.ca/health/story/2006/07/12/mollusc-fossil.html