 | |
 | |
| Идентификаторы | |
|---|---|
| ChemSpider | |
PubChem CID | |
| Характеристики | |
| C 18 H 10 N 2 O 4 | |
| Молярная масса | 318,288 г · моль -1 |
| Плотность | От 1,6 до 1,8 г / см 3 |
| Температура плавления | <-20 ° С (-4 ° F, 253 К) |
| Точка кипения | От 450 до 550 ° C (от 842 до 1022 ° F; от 723 до 823 K) |
Если не указано иное, данные приведены для материалов в их стандартном состоянии (при 25 ° C [77 ° F], 100 кПа). | |
| Ссылки на инфобоксы | |

{kind=link}
{kind=link}
Меланин ( / м ɛ л ə п ɪ п / ( слушать )![]() , от греческого : μέλας Мелас , «черный, темный») представляет собой общий термин для группы природных пигментов можно найти в большинстве организмов . Меланин производится через многоступенчатый химический процесс , известный как меланогенез , где окисление на аминокислоты тирозина следует полимеризация . Пигменты меланина производятся в специальной группе клеток, известной как меланоциты .
, от греческого : μέλας Мелас , «черный, темный») представляет собой общий термин для группы природных пигментов можно найти в большинстве организмов . Меланин производится через многоступенчатый химический процесс , известный как меланогенез , где окисление на аминокислоты тирозина следует полимеризация . Пигменты меланина производятся в специальной группе клеток, известной как меланоциты .
Существует пять основных типов меланина: эумеланин , феомеланин , нейромеланин , алломеланин и пиомеланин. [1] Наиболее распространенным типом является эумеланин, который бывает двух типов: коричневый эумеланин и черный эумеланин. Феомеланина является цистеином -производным , который содержит поли бензотиазин часть, которые в значительной степени ответственны за цвет красных волос , среди другой пигментации. Нейромеланин находится в головном мозге . Было проведено исследование его эффективности при лечении нейродегенеративных расстройств, таких как болезнь Паркинсона . [2] Алломеланин и пиомеланин - это два типа безазотистого меланина.
В коже человека меланогенез инициируется воздействием УФ-излучения , в результате чего кожа темнеет. Меланин - эффективный поглотитель света; пигмент способен рассеивать более 99,9% поглощенного УФ-излучения. [3] Из-за этого свойства считается, что меланин защищает клетки кожи от повреждения УФ-излучением , снижая риск истощения фолиевой кислоты и деградации дермы, и считается, что воздействие УФ-излучения связано с повышенным риском злокачественной меланомы , рака. меланоцитов (клеток меланина). Исследования показали меньшую заболеваемость раком кожи у людей с более концентрированным меланином, то есть более темным оттенком кожи . Однако связь между пигментацией кожи ифотозащита все еще не определена. [4]
Люди [ править ]
Этот раздел требует дополнительных ссылок для проверки . ( Март 2015 г. ) ( Узнайте, как и когда удалить этот шаблон сообщения ) |
У людей меланин является основным определяющим фактором цвета кожи . Он также содержится в волосах , пигментированные ткани , лежащие в основе радужной оболочки глаза, и бороздка vascularis из внутреннего уха . В головном мозге ткани с меланином включают мозговое вещество и нейроны, несущие пигмент, в областях ствола мозга , таких как голубое пятно . Это также происходит в сетчатых в надпочечниках . [5]
Меланина в коже вырабатывается меланоцитов , которые находятся в базальном слое из эпидермиса . Хотя в целом люди обладают одинаковой концентрацией меланоцитов в коже, у некоторых людей и этнических групп меланоциты производят различное количество меланина. У некоторых людей в организме очень мало или совсем отсутствует синтез меланина - состояние, известное как альбинизм . [6]
Поскольку меланин представляет собой совокупность более мелких компонентных молекул, существует множество различных типов меланина с разными пропорциями и схемами связывания этих компонентных молекул. И феомеланин, и эумеланин обнаруживаются в коже и волосах человека, но эумеланин является наиболее распространенным меланином у людей, а также формой, которая, скорее всего, будет дефицитной при альбинизме. [7]
Эумеланин [ править ]
Эумеланин полимеры уже давно мысль включает в себя многочисленный сшитый 5,6-дигидрокси индол ( DHI ) и 5,6-дигидроксииндол-2- карбоновой кислоты ( DHICA ) полимеры. [8]
Существует два типа эумеланина: коричневый эумеланин и черный эумеланин. Эти два типа эумеланина химически отличаются друг от друга характером полимерных связей. Небольшое количество черного эумеланина при отсутствии других пигментов вызывает седину. Небольшое количество коричневого эумеланина при отсутствии других пигментов вызывает желтые (светлые) волосы. По мере старения организм продолжает вырабатывать черный эумеланин, но перестает вырабатывать коричневый эумеланин, что приводит к появлению седых волос, характерных для пожилых людей. [9]
Феомеланин [ править ]
Феомеланины (или феомеланины) придают диапазон от желтоватого до красноватого цвета. [10] Феомеланины особенно сконцентрированы в губах, сосках, головке полового члена и влагалище. [11] Когда небольшое количество коричневого эумеланина в волосах, которое в противном случае привело бы к появлению светлых волос, смешивается с красным феомеланином, в результате появляются оранжевые волосы, которые обычно называют «красными» или «рыжими» волосами . Феомеланин также присутствует в коже, и, следовательно, у рыжих часто бывает более розоватый оттенок кожи.
С химической точки зрения феомеланины отличаются от эумеланинов тем, что структура олигомера включает в себя производимые бензотиазиновые и бензотиазольные звенья [12] вместо DHI и DHICA , когда присутствует аминокислота L-цистеин .
Трихромы [ править ]
Трихохромы (ранее называемые трихосидеринами) - это пигменты, производимые тем же метаболическим путем, что и эумеланины и феомеланины, но в отличие от этих молекул они имеют низкую молекулярную массу. Они встречаются в некоторых рыжих человеческих волосах. [13]
Нейромеланин [ править ]
Нейромеланин (НМ) - это темный нерастворимый полимерный пигмент, вырабатываемый в определенных популяциях катехоламинергических нейронов головного мозга. У людей самое большое количество ЯМ, которое присутствует в меньших количествах у других приматов и полностью отсутствует у многих других видов. [14] Биологическая функция остается неизвестной, хотя было показано, что человеческий НМ эффективно связывает переходные металлы, такие как железо, а также другие потенциально токсичные молекулы. Следовательно, он может играть решающую роль в апоптозе и связанной с ним болезни Паркинсона . [15]
Другие организмы [ править ]
Меланины выполняют очень разные роли и функции в разных организмах. Форма меланина составляет чернила, используемые многими головоногими моллюсками (см. Чернила головоногих ) в качестве защитного механизма от хищников. Меланины также защищают микроорганизмы, такие как бактерии и грибы, от стрессов, вызывающих повреждение клеток, таких как УФ-излучение от солнца и активные формы кислорода . Меланин также защищает от повреждений, вызванных высокими температурами, химическими воздействиями (такими как тяжелые металлы и окислители ) и биохимическими угрозами (такими как защита хозяина от вторгшихся микробов). [16] Следовательно, у многих патогенных микробов (например, у Cryptococcus neoformans, гриб) меланины, по-видимому, играют важную роль в вирулентности и патогенности , защищая микроб от иммунных ответов его хозяина . У беспозвоночных основным аспектом системы врожденной иммунной защиты от вторжения патогенов является меланин. Через несколько минут после заражения микроб инкапсулируется в меланине (меланизация), и считается, что образование побочных продуктов свободных радикалов во время образования этой капсулы помогает их убить. [17] Некоторые типы грибов, называемые радиотрофными грибами , по-видимому, могут использовать меланин в качестве фотосинтетического пигмента, который позволяет им улавливать гамма-лучи [18]и использовать эту энергию для роста. [19]
Более темные перья из птиц обязаны своим цветом меланина и менее легко разлагаются бактериями , чем непигментированных те или те , которые содержат каротиноиды пигменты. [20] Перья, содержащие меланин, также на 39% более устойчивы к истиранию, чем перья, которые не содержат, потому что гранулы меланина помогают заполнить пространство между кератиновыми прядями, которые образуют перья. [21] [22]Синтез феомеланина у птиц подразумевает потребление цистеина, полузаменимой аминокислоты, которая необходима для синтеза антиоксидантного глутатиона (GSH), но при избытке в рационе может быть токсичным. Действительно, хищные птицы, в рационе которых много белка, имеют окраску на основе феомеланина. [23]
Меланин также важен для пигментации млекопитающих . [24] Структура шерсти млекопитающих определяется геном агути, который регулирует распределение меланина. [25] [26] Механизмы действия гена были тщательно изучены на мышах, чтобы дать представление о разнообразии структур шерсти млекопитающих. [27]
Наблюдалось, что меланин у членистоногих откладывается слоями, создавая брэгговский отражатель с переменным показателем преломления. Когда масштаб этого рисунка совпадает с длиной волны видимого света, возникает структурная окраска , придающая ряду видов переливающийся цвет. [28]
Паукообразные - одна из немногих групп, в которых меланин обнаружить нелегко, хотя исследователи обнаружили данные, свидетельствующие о том, что пауки действительно производят меланин. [29]
Некоторые виды бабочек, в том числе лесная тигровая , превращают ресурсы в меланин, чтобы улучшить свою терморегуляцию. Поскольку у древесной тигровой бабочки есть популяции в большом диапазоне широт, было замечено, что более северные популяции показали более высокие темпы меланизации. Как у желтых, так и у белых самцов лесной тигровой моли, особи с большим количеством меланина имели повышенную способность улавливать тепло, но повышенный уровень хищничества из-за более слабого и менее эффективного апосематического сигнала. [30]
Растения [ править ]
Меланин, вырабатываемый растениями, иногда называют «катехоловые меланины», поскольку они могут давать катехол при плавлении с щелочами. Это часто наблюдается в ферментативном потемнении фруктов, таких как бананы. Меланин из скорлупы каштана можно использовать как антиоксидант и краситель. [31] Биосинтез включает окисление индол-5,6-хинона полифенолоксидазой тирозиназного типа из тирозина и катехоламинов, что приводит к образованию катехол-меланина. Несмотря на это, многие растения содержат соединения, подавляющие выработку меланинов. [32]
Биосинтетические пути [ править ]
Первый этап пути биосинтеза обоих eumelanins и pheomelanins является катализируемой с помощью тирозиназы . [33]
- Тирозин → ДОФА → Допахинон
Допахинон может соединяться с цистеином двумя путями с образованием бензотиазинов и феомеланинов.
- Допахинон + цистеин → 5-S-цистеинилдопа → промежуточный бензотиазин → феомеланин
- Допахинон + цистеин → 2-S-цистеинилдопа → промежуточный бензотиазин → феомеланин
Кроме того, допахинон может быть преобразован в лейкодопахром и пройти еще два пути к эумеланинам.
- Допахинон → лейкодопахром → допахром → 5,6-дигидроксииндол-2-карбоновая кислота → хинон → эумеланин
- Допахинон → лейкодопахром → допахром → 5,6-дигидроксииндол → хинон → эумеланин
Подробные метаболические пути можно найти в базе данных KEGG (см. Внешние ссылки ).
Микроскопический вид [ править ]
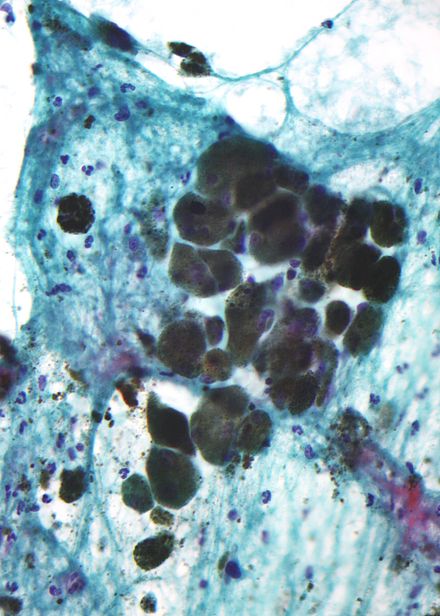
Меланин коричневый, непреломляющий и мелкозернистый с отдельными гранулами, имеющими диаметр менее 800 нанометров. Это отличает меланин от обычных пигментов , разлагающих кровь , которые крупнее, крупнее и преломлются, а их цвет варьируется от зеленого до желтого или красно-коричневого. В сильно пигментированных поражениях плотные агрегаты меланина могут скрывать гистологические детали. Разбавленный раствор перманганата калия - эффективный отбеливатель меланина. [34]
Генетические расстройства и болезненные состояния [ править ]
Существует около девяти типов глазно-кожного альбинизма , который в основном является аутосомно-рецессивным заболеванием. У некоторых этнических групп больше случаев различных форм. Например, наиболее распространенный тип, называемый кожно-кожным альбинизмом 2 типа (OCA2), особенно часто встречается у лиц чернокожего африканского происхождения. Это аутосомно-рецессивное заболевание, характеризующееся врожденным уменьшением или отсутствием пигмента меланина в коже, волосах и глазах. По оценкам, частота OCA2 среди афроамериканцев составляет 1 из 10 000, что контрастирует с частотой 1 из 36 000 среди белых американцев. [35] В некоторых африканских странах частота заболевания еще выше - от 1 из 2 000 до 1 из 5 000.[36] Другая форма альбинизма, «желтый глазокожный альбинизм», по-видимому, более распространена среди амишей , которые имеют преимущественно швейцарское и немецкое происхождение. У людей с этим вариантом IB обычно белые волосы и кожа при рождении, но в младенчестве быстро развивается нормальная пигментация кожи. [36]
Глазной альбинизм влияет не только на пигментацию глаз, но и на остроту зрения. Люди с альбинизмом обычно плохо проходят тесты, в диапазоне от 20/60 до 20/400. Кроме того, две формы альбинизма, примерно у 1 из 2700 наиболее распространенных среди людей пуэрториканского происхождения, связаны со смертностью, не считая смертей, связанных с меланомой.
Связь между альбинизмом и глухотой хорошо известна, но плохо изучена. В 1859 году трактат О происхождении видов , Чарльз Дарвин заметил , что «кошки , которые полностью белые и голубые глаза , как правило , глухие». [37] У людей гипопигментация и глухота встречаются вместе в редком синдроме Ваарденбурга , который преимущественно наблюдается у хопи в Северной Америке . [38] Заболеваемость альбинизмом у индейцев хопи оценивается примерно в 1 из 200 человек. Подобные образцы альбинизма и глухоты были обнаружены у других млекопитающих, включая собак и грызунов. Однако недостаток меланина как таковогопо-видимому, не несет прямой ответственности за глухоту, связанную с гипопигментацией, поскольку у большинства людей, лишенных ферментов, необходимых для синтеза меланина, слуховая функция нормальная. [39] Вместо этого отсутствие меланоцитов в сосудистой полоске внутреннего уха приводит к нарушению улитки , [40] хотя, почему это так, до конца не выяснено.
При болезни Паркинсона, расстройстве, влияющем на нейромоторное функционирование, наблюдается снижение нейромеланина в черной субстанции и голубом пятне как следствие специфического выпадения дофаминергических и норадренергических пигментированных нейронов. Это приводит к снижению синтеза дофамина и норэпинефрина . Хотя о корреляции между расой и уровнем нейромеланина в черной субстанции не сообщалось, значительно более низкая частота болезни Паркинсона у чернокожих, чем у белых, «подтолкнула некоторых к предположению, что кожный меланин может каким-то образом служить для защиты нейромеланина в субстанции. черная от внешних токсинов ". [41]
Помимо дефицита меланина, молекулярная масса полимера меланина может быть уменьшена различными факторами, такими как окислительный стресс, воздействие света, нарушение его связи с белками меланосомного матрикса , изменения pH или локальные концентрации ионов металлов. Было предложено снизить молекулярную массу или степень полимеризации глазного меланина , чтобы превратить обычный антиоксидантный полимер в прооксидант . Предполагается, что в своем прооксидантном состоянии меланин участвует в возникновении и прогрессировании дегенерации желтого пятна и меланомы . [42] Расагилин, важный монотерапевтический препарат при болезни Паркинсона, обладает свойствами связывания меланина и свойствами уменьшения опухоли меланомы. [43]
Однако более высокие уровни эумеланина также могут быть недостатком, помимо более высокой предрасположенности к дефициту витамина D. Темная кожа является осложняющим фактором при лазерном удалении винных пятен . Как правило, лазеры эффективны для лечения белой кожи и менее успешно удаляют пятна портвейна у людей азиатского или африканского происхождения. Более высокие концентрации меланина у темнокожих людей просто рассеивают и поглощают лазерное излучение, препятствуя поглощению света тканью-мишенью. Аналогичным образом меланин может осложнить лазерное лечение других дерматологических заболеваний у людей с более темной кожей.
Веснушки и родинки образуются там, где в коже имеется локальная концентрация меланина. Они часто ассоциируются с бледной кожей.
Никотин имеет сродство с меланинсодержащими тканями из-за его функции-предшественника в синтезе меланина или необратимом связывании меланина. Было высказано предположение, что это лежит в основе повышенной никотиновой зависимости и более низких показателей отказа от курения у лиц с более темным пигментом. [44]
Адаптация человека [ править ]
Физиология [ править ]
Меланоциты вставляют гранулы меланина в специализированные клеточные пузырьки, называемые меланосомами . Затем они переносятся в клетки кератиноцитов эпидермиса человека . Меланосомы в каждой клетке-реципиенте накапливаются наверху ядра клетки , где они защищают ядерную ДНК от мутаций, вызванных ионизирующим излучением ультрафиолетовых лучей солнца . В общем, люди, чьи предки долгое время жили в регионах земного шара вблизи экватора.в их коже содержится большее количество эумеланина. Это делает их кожу коричневой или черной и защищает их от сильного воздействия солнца, которое чаще приводит к меланомам у людей со светлой кожей. [45]
Не все эффекты пигментации благоприятны. Пигментация увеличивает тепловую нагрузку в жарком климате, и темнокожие люди поглощают на 30% больше тепла от солнечного света, чем очень светлокожие люди, хотя этот фактор может быть компенсирован более обильным потоотделением. В холодном климате темная кожа приводит к большей потере тепла за счет излучения. Пигментация также препятствует синтезу витамина D , поэтому в районах с плохим питанием дети с более темной кожей более подвержены рахиту, чем дети со светлой кожей. Поскольку пигментация кажется не совсем полезной для жизни в тропиках, были выдвинуты другие гипотезы о ее биологическом значении, например, вторичное явление, вызванное адаптацией к паразитам и тропическим болезням. [46]
Эволюционное происхождение [ править ]
Ранние люди приобрели темный цвет кожи около 1,2 миллиона лет назад в результате адаптации к потере волос на теле, которая усилила воздействие ультрафиолетового излучения. До появления безволосости у древних людей под шерстью была достаточно светлая кожа, похожая на ту, что есть у других приматов . [47] Самые последние научные данные показывают, что анатомически современные люди эволюционировали в Африке между 200 000 и 100 000 лет [48], а затем заселили остальной мир в результате одной миграции между 80 000 и 50 000 лет назад, в некоторых областях скрещиваясь с некоторыми архаическими человеческий вид ( неандертальцы ,Денисовцы и, возможно, другие). [49] Кажется вероятным, что у первых современных людей было относительно большое количество меланоцитов, продуцирующих эумеланин, производящих более темную кожу, как у коренных жителей Африки сегодня. Поскольку некоторые из этих коренных народов мигрировали и поселились в регионах Азии и Европы , селективное давление на производство эумеланина уменьшилось в климате, где солнечное излучение было менее интенсивным. В конечном итоге это привело к нынешнему диапазону цвета кожи человека. Из двух распространенных вариантов генов, связанных с бледной кожей человека, Mc1r , по-видимому, не подвергся положительной селекции [50], в то время как SLC24A5 подвергся положительной селекции.[51]
Эффекты [ править ]
Как и люди, мигрировавшие на север, люди со светлой кожей, мигрирующие к экватору, акклиматизируются к гораздо более сильному солнечному излучению. Природа выбирает меньше меланина, когда ультрафиолетовое излучение слабое. Кожа большинства людей темнеет под воздействием ультрафиолета, что дает им дополнительную защиту, когда это необходимо. Это физиологическая цель загара . Темнокожие люди, которые производят больше эумеланина, защищающего кожу, имеют лучшую защиту от солнечных ожогов и развития меланомы, потенциально смертельной формы рака кожи, а также других проблем со здоровьем, связанных с воздействием сильного солнечного излучения , включая фотодеградацию некоторых витаминов, таких как рибофлавины, каротиноиды , токоферол и фолиевая кислота . [52] Некоторые северо-западные европейцы существенно утратили способность загорать в результате ослабления естественного отбора. Их кожа горит и шелушится, а не загорает. Это связано с тем, что они производят дефектную форму кожного белка Mc1r (рецептор меланокортина-1), который необходим для производства меланина. В тропических и субтропических средах они явно находятся в невыгодном положении. Они не только страдают от дискомфорта, связанного с легкостью обжигания, но и подвергаются гораздо более высокому риску рака кожи; то же самое и с альбиносами. [53]
Меланин в глазах, в радужке и сосудистой оболочке помогает защитить их от ультрафиолета и высокочастотного видимого света ; люди с серыми, голубыми и зелеными глазами больше подвержены риску заболеваний глаз, связанных с солнцем. Кроме того, линза глаза желтеет с возрастом, обеспечивая дополнительную защиту. Однако с возрастом хрусталик также становится более жестким, теряя большую часть своей аккомодации - способности изменять форму, чтобы фокусироваться с дальнего на ближний - ущерб, вероятно, из-за сшивания белков, вызванного воздействием УФ-излучения.
Недавние исследования показывают, что меланин может выполнять не только фотозащитную функцию, но и не только для защиты от света. [54] Меланин способен эффективно хелатировать ионы металлов через свои карбоксилатные и фенольные гидроксильные группы, во многих случаях гораздо более эффективно, чем мощный хелатирующий лиганд этилендиаминтетраацетат (ЭДТА). Таким образом, он может служить для изоляции потенциально токсичных ионов металлов, защищая остальную часть клетки. Эта гипотеза подтверждается тем фактом, что потеря нейромеланина, наблюдаемая при болезни Паркинсона, сопровождается увеличением уровня железа в головном мозге.
Физические свойства и технологические приложения [ править ]
Существуют доказательства в поддержку высокосшитого гетерополимера, ковалентно связанного с матриксными каркасными меланопротеинами . [55] Было высказано предположение, что способность меланина действовать как антиоксидант прямо пропорциональна его степени полимеризации или молекулярной массе . [56] Неоптимальные условия для эффективной полимеризации мономеров меланина могут привести к образованию прооксидантного меланина с более низкой молекулярной массой, который является причиной и развитием дегенерации желтого пятна и меланомы . [57] Сигнальные пути, которыеактивируют меланизации в пигментном эпителии сетчатки (RPE) , также могут быть вовлечены в понижающей из стержня наружного сегмента фагоцитоза с помощью ПЭС. Это явление частично объясняется сохранением фовеа при дегенерации желтого пятна . [58]
См. Также [ править ]
- Альбинос
- Альбинизм в биологии
- Синдром Гричелли, синдром , характеризующийся гипопигментацией.
- Цвет кожи человека
- Теория меланина
- Меланизм
- Меланогенез , производство меланина
- Риски и преимущества пребывания на солнце
- Витамин Д
- Отбеливание кожи
- Феруловая кислота
Ссылки [ править ]
- ^ Цао, Вэй; Чжоу, Сюхао; McCallum, Naneki C .; Ху, Цзыин; Ни, Цин Чжэ; Капур, Уткарш; Heil, Christian M .; Cay, Kristine S .; Занд, Тара; Мантанона, Алекс Дж .; Джаяраман, Арти (2021-02-09). «Раскрытие структуры и функции меланина посредством синтеза» . Журнал Американского химического общества . DOI : 10.1021 / jacs.0c12322 . ISSN 0002-7863 .
- ^ Хайнинг, Роберт Л .; Ахат-Мендес, Синди (март 2017 г.). «Нейромеланин, одна из самых недооцененных молекул в современной медицине, не является зрителем» . Исследование нейронной регенерации . 12 (3): 372–375. DOI : 10.4103 / 1673-5374.202928 . PMC 5399705 . PMID 28469642 .
- Перейти ↑ Meredith P, Riesz J (2004). «Квантовые выходы радиационной релаксации синтетического эумеланина». Фотохимия и фотобиология . 79 (2): 211–6. arXiv : cond-mat / 0312277 . DOI : 10.1111 / j.1751-1097.2004.tb00012.x . PMID 15068035 . S2CID 222101966 .
- Перейти ↑ Brenner M, Hearing VJ (2008). «Защитная роль меланина против УФ-повреждений кожи человека» . Фотохимия и фотобиология . 84 (3): 539–49. DOI : 10.1111 / j.1751-1097.2007.00226.x . PMC 2671032 . PMID 18435612 .
- Перейти ↑ Solano, F. (2014). «Меланины: пигменты кожи и многое другое - типы, структурные модели, биологические функции и пути образования» . Новый научный журнал . 2014 : 1–28. DOI : 10.1155 / 2014/498276 .
- ^ Цихорек, Мирослава; Вачульска, Малгожата; Стасевич, Анета; Тиминьская, Агата (20 февраля 2013 г.). «Меланоциты кожи: биология и развитие» . Успехи дерматологии и аллергологии . 30 (1): 30–41. DOI : 10,5114 / pdia.2013.33376 . PMC 3834696 . PMID 24278043 .
- ^ «Глазокожный альбинизм» . Домашний справочник по генетике . Проверено 25 сентября 2017 .
- ^ Мередит, Пол; Сарна, Тадеуш (01.12.2006). «Физико-химические свойства эумеланина». Исследование пигментных клеток . 19 (6): 572–594. DOI : 10.1111 / j.1600-0749.2006.00345.x . PMID 17083485 .
- ^ Ито, S .; Вакамацу, К. (декабрь 2011 г.). «Разнообразие пигментации волос человека, изученное с помощью химического анализа эумеланина и феомеланина». Журнал Европейской академии дерматологии и венерологии: JEADV . 25 (12): 1369–1380. DOI : 10.1111 / j.1468-3083.2011.04278.x . ISSN 1468-3083 . PMID 22077870 . S2CID 5121042 .
- ^ Slominski A, Тобин DJ, Shibahara S, Wortsman J (октябрь 2004). «Пигментация меланина в коже млекопитающих и ее гормональная регуляция» . Физиологические обзоры . 84 (4): 1155–228. DOI : 10.1152 / Physrev.00044.2003 . PMID 15383650 . S2CID 21168932 .
- ^ «феомеланин» . База данных MetaCyc Metabolic Pathway . 2010 г.[ требуется полная ссылка ]
- ^ Greco G, Panzella L, L Verotta, д'Искья M Наполитано A (апрель 2011). «Раскрытие структуры феомеланина рыжих волос человека: бензотиазолилтиазинодигидроизохинолины как ключевые строительные блоки». Журнал натуральных продуктов . 74 (4): 675–82. DOI : 10.1021 / np100740n . PMID 21341762 .
- ^ Prota, G .; Серл, AG (1978). «Биохимические участки действия генов меланогенеза у млекопитающих» . Annales de Genétique et de Sélection Animale . 10 (1): 1–8. DOI : 10.1186 / 1297-9686-10-1-1 . PMC 2757330 . PMID 22896083 .
- ^ Федоров Н, Tribl F, G Холлидей, Герлах M, Riederer P, Double KL (2005). «Нейромеланин в нейронах дофамина человека: сравнение с периферическими меланинами и отношение к болезни Паркинсона». Prog Neurobiol . 75 (2): 109–124. DOI : 10.1016 / j.pneurobio.2005.02.001 . PMID 15784302 . S2CID 503902 .
- ^ Двойной KL (2006). «Функциональные эффекты нейромеланина и синтетического меланина в модельных системах». J Neural Transm . 113 (6): 751–756. DOI : 10.1007 / s00702-006-0450-5 . PMID 16755379 . S2CID 23096297 .
- ^ Гамильтон AJ, Гомес BL (март 2002). «Меланины в грибковых возбудителях» . Журнал медицинской микробиологии . 51 (3): 189–91. DOI : 10.1099 / 0022-1317-51-3-189 . PMID 11871612 .
- ^ Cerenius L, Söderhäll K (апрель 2004). «Система активации пропенолоксидазы у беспозвоночных». Иммунологические обзоры . 198 : 116–26. DOI : 10.1111 / j.0105-2896.2004.00116.x . PMID 15199959 . S2CID 10614298 .
- Перейти ↑ Castelvecchi, Davide (26 мая 2007 г.). «Темная сила: пигмент, кажется, находит хорошее применение радиации». Новости науки . 171 (21): 325. DOI : 10.1002 / scin.2007.5591712106 .
- ^ Дадачева Э., Брайан Р.А., Хуанг Х и др. (2007). «Ионизирующее излучение изменяет электронные свойства меланина и усиливает рост меланизированных грибов» . PLOS ONE . 2 (5): e457. Bibcode : 2007PLoSO ... 2..457D . DOI : 10.1371 / journal.pone.0000457 . PMC 1866175 . PMID 17520016 .
- ^ Gunderson, Alex R .; Кадр, Алисия М .; Swaddle, Джон П .; Форсайт, Марк Х. (01.09.2008). «Устойчивость меланизированных перьев к бактериальной деградации: неужели все такое черно-белое?». Журнал биологии птиц . 39 (5): 539–545. DOI : 10.1111 / j.0908-8857.2008.04413.x .
- ^ Бонсер, Ричард ХК (1995). «Меланин и сопротивление истиранию перьев» . Кондор . 97 (2): 590–591. DOI : 10.2307 / 1369048 . JSTOR 1369048 .
- ^ Гальбан, Исмаэль; Солано, Франциско (2016-04-08). "Меланины покровов птиц: биосинтез, формы, функции и эволюция" . Международный журнал молекулярных наук . 17 (4): 520. DOI : 10,3390 / ijms17040520 . PMC 4848976 . PMID 27070583 .
- ^ Родригес-Мартинес, Сол; Гальван, Исмаэль (2020). «Окраска оперения молоди на основе феомеланина чаще развивалась у хищных видов». Ибис . 162 (1): 238–244. DOI : 10.1111 / ibi.12770 . hdl : 10261/207451 . ISSN 1474-919X .
- ^ Джимбоу, K; Кеведо WC, младший; Фитцпатрик, ТБ; Сабо, Г. (июль 1976 г.). «Некоторые аспекты биологии меланина: 1950–1975» . Журнал следственной дерматологии . 67 (1): 72–89. DOI : 10.1111 / 1523-1747.ep12512500 . PMID 819593 .
- ^ Meneely, Филипп (2014). Генетический анализ: гены, геномы и сети у эукариот . Издательство Оксфордского университета. ISBN 9780199681266.
- ^ Гриффитс, Энтони JF; Миллер, Джеффри Х .; Судзуки, Дэвид Т .; Левонтин, Ричард С .; Гелбарт, Уильям М. (2000). «Взаимодействие генов в окрасе шерсти млекопитающих» . Цитировать журнал требует
|journal=( помощь ) - ^ Миллар, ЮВ; Миллер, МВт; Стивенс, Мэн; Барш, GS (октябрь 1995 г.). «Экспрессия и трансгенные исследования гена агути у мышей дают представление о механизмах, с помощью которых генерируются образцы окраски шерсти млекопитающих». Развитие . 121 (10): 3223–3232. PMID 7588057 .
- Перейти ↑ Neville, AC (2012). Биология кутикулы членистоногих . Springer Science & Business Media. ISBN 9783642809101.
- ^ Hsiung, B.-K .; Блэкледж, TA; Шоуки, доктор медицины (2015). «У пауков все-таки есть меланин» . Журнал экспериментальной биологии . 218 (22): 3632–3635. DOI : 10,1242 / jeb.128801 . PMID 26449977 .
- ^ Hegna, Роберт Х .; Нокелайнен, Осси; Hegna, Jonathan R .; Маппс, Джоанна (2013). «Дрожь или дрожь: усиление меланизации способствует терморегуляции, но снижает эффективность предупреждающих сигналов у древесной тигровой бабочки» . Proc. R. Soc. B . 280 (1755): 20122812. DOI : 10.1098 / rspb.2012.2812 . PMC 3574392 . PMID 23363631 .
- ^ Яо, Цзэн-Ю; Ци, Цзянь-Хуа (22 апреля 2016 г.). «Сравнение антиоксидантной активности фракций меланина из скорлупы каштана» . Молекулы . 21 (4): 487. DOI : 10.3390 / modules21040487 . PMC 6273334 . PMID 27110763 .
- ^ Kim, Y.-J .; Уяма, Х. (15 мая 2005 г.). «Ингибиторы тирозиназы из природных и синтетических источников: структура, механизм ингибирования и перспективы на будущее». Клеточные и молекулярные науки о жизни . 62 (15): 1707–1723. DOI : 10.1007 / s00018-005-5054-у . PMID 15968468 . S2CID 8280251 .
- ^ Зайди, Камаль Uddin; Али, Аиша С .; Али, Шарик А .; Нааз, Ишрат (2014). «Микробные тирозиназы: многообещающие ферменты для фармацевтической, пищевой и биотехнологической промышленности» . Международное исследование биохимии . 2014 : 1–16 (см. Рис. 3). DOI : 10.1155 / 2014/854687 . PMC 4033337 . PMID 24895537 .
- ^ "Меланин" . pubchem.ncbi.nlm.nih.gov . Проверено 25 сентября 2017 .
- ^ "Глазокожный альбинизм" . Архив с оригинала на 23 декабря 2008 года.
- ^ a b «Глазные проявления альбинизма: история вопроса, патофизиология, эпидемиология» . 18 июня 2018 г. - через eMedicine. Цитировать журнал требует
|journal=( помощь ) - ^ «Причины изменчивости» . Архивировано из оригинального 23 сентября 2006 года . Проверено 18 сентября 2006 года .
- ^ EntrezGene 300700
- ^ EntrezGene 606933
- ^ Кабель J, D Huszar, Йениш R, Стальной КП (февраль 1994 г.). «Влияние мутаций в локусе W (c-kit) на пигментацию внутреннего уха и функцию у мышей». Исследование пигментных клеток . 7 (1): 17–32. DOI : 10.1111 / j.1600-0749.1994.tb00015.x . PMID 7521050 .
- ^ "Болезнь тела Леви" . Архивировано из оригинала 21 июля 2009 года.
- ^ Meyskens FL, Farmer P, Fruehauf JP (июнь 2001). «Редокс-регуляция в меланоцитах и меланоме человека» (PDF) . Исследование пигментных клеток . 14 (3): 148–54. DOI : 10.1034 / j.1600-0749.2001.140303.x . PMID 11434561 .
- ^ Meier-Davis SR, Dines K, Arjmand FM и др. (Декабрь 2012 г.). «Сравнение перорального и трансдермального введения мезилата разагилина на рост опухоли меланомы человека in vivo». Кожная и глазная токсикология . 31 (4): 312–7. DOI : 10.3109 / 15569527.2012.676119 . PMID 22515841 . S2CID 30344869 .
- ^ King G, Yerger VB, Whembolua GL, Bendel RB, Kittles R, Moolchan ET (июнь 2009). «Связь между факультативным меланином и употреблением табака среди афроамериканцев». Фармакология, биохимия и поведение . 92 (4): 589–96. DOI : 10.1016 / j.pbb.2009.02.011 . PMID 19268687 . S2CID 3070838 .
- ^ "Изменение цвета кожи человека" . Программа "Происхождение человека" Смитсоновского института . 2012-06-20 . Проверено 24 августа 2019 .
- ^ Берт-Джонс, Дж. (2010), «Основная пигментация, пигментация человека и реакция на воздействие солнца», Тони Бернс; Стивен Бретнах; Нил Кокс; Кристофер Гриффитс (ред.), Учебник дерматологии Рока , 3 (8-е изд.), Wiley-Blackwell, стр. 58,9, ISBN 978-1-4051-6169-5
- ^ Уэйд, Николас (2003-08-19). «Почему люди и их мех разошлись» . Нью-Йорк Таймс . ISSN 0362-4331 . Проверено 24 августа 2019 .
- ^ Тишкофф С.А., Рид Ф.А., Фридлендер Ф.Р. и др. (Май 2009 г.). «Генетическая структура и история африканцев и афроамериканцев» . Наука . 324 (5930): 1035–44. Bibcode : 2009Sci ... 324.1035T . DOI : 10.1126 / science.1172257 . PMC 2947357 . PMID 19407144 .
- ^ "Единственная миграция из Африки заселила мир, исследования показывают" . Нью-Йорк Таймс . 2016-09-22.
- ^ Хардинг Р.М., Хили Э., Рэй А.Дж. и др. (Апрель 2000 г.). «Доказательства переменных избирательных давлений на MC1R» . Американский журнал генетики человека . 66 (4): 1351–61. DOI : 10.1086 / 302863 . PMC 1288200 . PMID 10733465 .
- ^ Lamason RL, Mohideen MA, Mest JR и др. (Декабрь 2005 г.). «SLC24A5, предполагаемый катионообменник, влияет на пигментацию у рыбок данио и людей». Наука . 310 (5755): 1782–6. Bibcode : 2005Sci ... 310.1782L . DOI : 10.1126 / science.1116238 . PMID 16357253 . S2CID 2245002 .
- ^ Яблонски, Нина Г .; Чаплин, Джордж (11.05.2010). «Пигментация кожи человека как адаптация к УФ-излучению» . Труды Национальной академии наук . 107 (Приложение 2): 8962–8968. Bibcode : 2010PNAS..107.8962J . DOI : 10.1073 / pnas.0914628107 . PMC 3024016 . PMID 20445093 .
- ^ https://www2.palomar.edu/anthro/adapt/adapt_4.htm
- Перейти ↑ Liu Y, Hong L, Kempf VR, Wakamatsu K, Ito S, Simon JD (июнь 2004 г.). «Ионный обмен и адсорбция Fe (III) меланином сепии». Исследование пигментных клеток . 17 (3): 262–9. DOI : 10.1111 / j.1600-0749.2004.00140.x . PMID 15140071 .
- ^ Donatien PD, Orlow SJ (август 1995). «Взаимодействие меланосомных белков с меланином». Европейский журнал биохимии . 232 (1): 159–64. DOI : 10.1111 / j.1432-1033.1995.tb20794.x . PMID 7556145 .
- ^ Sarangarajan R, Apte SP (2005). «Агрегация и полимеризация меланина: возможные последствия возрастной дегенерации желтого пятна». Офтальмологические исследования . 37 (3): 136–41. DOI : 10.1159 / 000085533 . PMID 15867475 . S2CID 27499198 .
- ^ Meyskens FL, Farmer PJ, Anton-Калвер H (апрель 2004). «Этиологический патогенез меланомы: объединяющая гипотеза отсутствующего риска» (PDF) . Клинические исследования рака . 10 (8): 2581–3. DOI : 10.1158 / 1078-0432.ccr-03-0638 . PMID 15102657 . S2CID 26079375 .
- ^ Sarangarajan R, Apte SP (2005). «Меланизация и фагоцитоз: последствия для возрастной дегенерации желтого пятна». Молекулярное зрение . 11 : 482–90. PMID 16030499 .
Внешние ссылки [ править ]
| Поищите меланин в Викисловаре, бесплатном словаре. |
| Найдите форму лейко в Викисловаре, бесплатном словаре. |
- «Спектр поглощения меланина» . Департамент компьютерных наук и технологий .
- «Метаболизм тирозина - контрольный путь» . Киотская энциклопедия генов и геномов .
- «Меланогенез - эталонный путь» . Киотская энциклопедия генов и геномов .